Autor: Peter Pančík
Publikované dňa:
Citácia: PANČÍK, Peter. 2016. Biopedia.sk: Regulácia bunkového cyklu. [cit. 2024-04-19]. Dostupné na internete: <https://biopedia.sk/bunka/regulacia-bunkoveho-cyklu>.
Pre správne fungovanie bunky a organizmu ako celku musí byť delenie buniek prísne kontrolované a regulované. Regulácia bunkového cyklu je zabezpečovaná bielkovinami, ktoré môžu byť dvojakého pôvodu:
- Exogénne regulátory sú mimobunkového pôvodu. Pre bunku sa teda jedná o reakciu na vonkajšie podmienky, ktoré predstavujú spravidla prvotný signál pre spustenie kaskády (dráhy) intracelulárnych biochemických reakcií. Patria sem rôzne hormóny a látky vylučované do tkanivového moku inými bunkami.
- Endogénne regulátory produkujú vlastné bunky podľa stavu bunkového cyklu. Patria sem hlavne bielkoviny kódované protoonkogénmi a tumor-supresorovými génmi.
Endogénna regulácia link
Protoonkogény a tumor-supresorové gény tvoria skupinu génov, ktoré kódujú špecifické regulačné bielkoviny bunkového cyklu. Ich mutáciou dochádza k deregulácii bunkového cyklu, nekontrolovateľnému deleniu buniek a rozvoju karcinogenézy.
Podstatou kontroly bunkového cyklu je kontrola integrity genetickej informácie na konci každej (!) fázy bunkového cyklu, kde sa nachádzajú tzv. kontrolné uzly (angl. checkpoint). Produkty protoonkogénov sú bielkoviny, ktoré posúvajú bunkový cyklus do ďalšej fázy, čím zároveň inhibujú aj bunkovú diferenciáciu. Patria sem cyklíny a cyklín-závislé kinázy (ďalej len CDK, angl. cyclin-dependent kinase). Produkty tumor-supresorových génov pôsobia opačne, tzn. inhibujú bunkový cyklus (bunkové delenie) a podporujú bunkovú diferenciáciu. Patria sem napr. produkty génov p53 alebo rb1.
Princíp regulácie bunkového cyklu nie je vôbec taký zložitý, ako by sa na prvý pohľad mohlo zdať. Kináza je všeobecné pomenovanie enzýmu, ktorý katalyzuje fosforyláciu substrátu (inej bielkoviny). Fosforylovaná bielkovina má mierne odlišné vlastnosti a priestorovú štruktúru ako jej nefosforylovaná forma, pričom táto zmena stavu predstavuje signál rozoznávaný ďalšími enzýmami, čím táto špecifická "informácia" postupuje po signálnej dráhe ďalej. CDK sú enzýmy, ktorých úlohou je fosforylovať cieľový substrát, ktorý potom predstavuje signál rozoznávaný radou ďalších molekúl a končí prechodom do ďalšej fázy bunkového cyklu (G1/S, S/G2, G2/M). Sami však fosforyláciu nezačnú, ale potrebujú na to väzbu s iným proteínom - cyklínom (preto cyklín-dependetné alebo cyklín-závislé kinázy). Práve cyklín určuje v komplexe so špecifickou CDK, ktorá cieľová molekula substrátu bude fosforylovaná. Pre každú fázu bunkového cyklu sú tak v bunke prítomné iné cyklíny. Na konci každej fázy sa cyklíny typické pre tú-ktorú fázu degradujú, aby nerušili lineárny priebeh bunkového cyklu. Produkty tumor-supresorových génov pôsobia opačne. Naviazaním sa na komplexy cyklín-CDK alebo aj samotné cyklíny spôsobujú ich zablokovanie, cieľový substrát nemôže byť fosforylovaný a bunkový cyklus je zablokovaný na čas potrebný k oprave poškodenia DNA. Ak bunka nie je schopná opraviť poškodenie, nastane apoptóza.
| protoonkogény | tumor-supresorové gény | |
| bunkový cyklus | stimulácia | inhibícia |
| diferenciácia | inhibícia | stimulácia |
| zástupcovia | cyklíny, CDK | p53, rb1 |
Kontrolné uzly:
- G1/S prechod - hlavný kontrolný uzol; kontroluje sa stav DNA, aby nedošlo k replikácii poškodenej DNA
- S/G2 prechod - kontroluje sa, či replikácia DNA prebehla bez chýb a prípadné chyby sa opravia
- G2/M prechod - ak sú všetky podmienky splnené a je dostatok enzýmov, bunka prejde do M-fázy
- mitotický kontrolný uzol - v metafáze sa kontroluje, či sú všetky chromozómy pripojené na deliace vretienko
Apoptóza link
V roku 2002 dostali S. Brenner, R. Horvitz, J. Sulston Nobelovu cenu za ich objavy súvisiace s genetickou reguláciou vývoja orgánov a programovanej smrti bunky. Ako už bolo skôr spomínané, ak reparačný aparát bunky zistí v niektorom z kontrolných uzlov nevratné poškodenie DNA, v bunke sa spustí kaskáda biochemických reakcií vedúca k tzv. programovanej bunkovej smrti - apoptóze. Zabraňuje sa tým malígnej transformácii poškodenej bunky, ktorá by mohla ohroziť celý organizmus. Apoptózou sú eliminované aj bunky napadnuté vírusmi, aby sa zabránilo ich rozmnožovaniu. Apoptóza zohráva úlohu aj pri embryogenéze a nidácii vajíčka. V navodení apoptózy majú významnú úlohu mitochondrie a endoplazmatické retikulum. Apoptóza je normálny fyziologický proces a prebieha v organizme bez zápalovej reakcie.
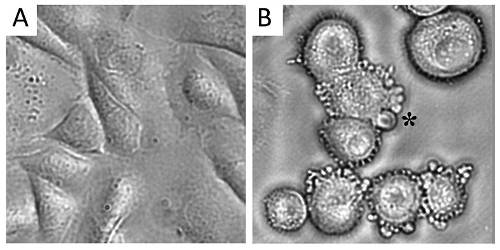
Bunky, ktoré podliehajú apoptóze, sú typické početnými pľuzgierikmi na ich povrchu, čo značí, že sa bunka rozpadáva.
Totipotencia a diferenciácia link
Každá bunka organizmu vzniká delením už existujúcej bunky. Keby sme sa pozerali retrospektívne na delenie buniek organizmu, došli by sme až k zygote. Vo všeobecnosti každá somatická bunka organizmu má rovnakú genetickú informáciu ako zygota. Prvé embryonálne bunky vzniknuté delením zygoty sú totipotentné. Totipotencia je vlastnosť, akýsi potenciál, že z danej bunky môže vzniknúť kompletný mnohobunkový organizmus.
Hoci sú dcérske bunky geneticky zhodné s materskou bunkou, u mnohobunkového organizmu postupne dochádza k zmene ich tvaru a vlastností. Hovoríme, že bunky sa postupne morfologicky diferencujú a funkčne špecializujú. Príčinou toho je aktívne prepisovanie len časti genetickej informácie. Regulácia tohto procesu predstavuje nesmierne zložitý komplex makromolekulových interakcií.
Proces bunkovej diferenciácie je u živočíšnych buniek v podstate nevratný a diferencované bunky nie je možné spätne dediferencovať, tzn. priviesť ich k prvotnému štádiu zygoty a vytvoriť z nich nový organizmus. U rastlinných buniek toto do istej miery možné je (príkadom sú kalusové kultúry).
Bunky ďalších generácií teda strácajú totipotenciu, pretože si zo sebou už nesú akési "nastavenia" rodičovskej bunkovej línie. Stále to však môžu byť bunky pluripotentné alebo multipotentné, z ktorých môže vzniknúť pomerne široký repertoár bunkových typov, nie však už celý mnohobunkový organizmus. U živočíšnych buniek spravidla existujú v tkanivách "zásoby" menej diferencovaných buniek, ktoré v prípade potreby svojim delením dopĺňajú stavy špecializovaných tkanív. Napokon najviac špecializovanými bunkami sú unipotentné bunky, ktoré dokážu dať vznik len bunkovému typu ako sú ony samé.
Faktory ovplyvňujúce diferenciáciu sú:
- vzájomný vplyv jadra a cytoplazmy
- vonkajšie prostredie
- embryonálna indukcia - vzájomný vplyv buniek v zárodku
- kontaktná inhibícia - vzájomný dotyk buniek zamedzuje ich rast a delenie
- pôsobenie signálnych molekúl, hormónov