Autor: Peter Pančík
Publikované dňa:
Citácia: PANČÍK, Peter. 2016. Biopedia.sk: Genetika prokaryot. [cit. 2024-04-20]. Dostupné na internete: <https://biopedia.sk/genetika/genetika-prokaryot>.
Prokaryotické organizmy nemožno geneticky študovať tak ako eukaryotické predovšetkým z toho dôvodu, že majú len jeden chromozóm, a teda sú kvázi po celý život v haploidnom, resp. monoploidnom stave. Teda komplementačný, segregačný a rekombinačný test u nich nepripadá do úvahy. Napriek tomu existujú možnosti, ako do prokaryotickej bunky vniesť DNA, pričom sa tieto procesy môžu využiť aj na mapovanie prokaryotických génov.
Genetický materiál sa medzi baktériami môže prenášať 3 základnými procesmi: konjugáciou, transdukciou a transformáciou. Každý z týchto procesov zahŕňa jednosmerný prenos DNA z jednej bunky do druhej. Rozlišujú sa podľa citlivosti k nukleázam (prítomných v kultivačnom médiu) a podľa toho, či sa na úspešný prenos DNA vyžaduje bunkový kontakt alebo nie.
| proces | citlivosť k nukleázam | bunkový kontakt |
| konjugácia | nie | áno |
| transdukcia | nie | nie |
| transformácia | áno | nie |
Auxotrofné mutanty a kultivačné médiá link
Základom štúdia spomenutých troch dejov je využitie bakteriálnych kmeňov - auxotrofných mutantov, ktorým chýbajú alebo sú mutované gény pre syntézu esenciálnych látok (aminokyseliny, dusíkaté bázy). Tieto zlúčeniny sú však bunky schopné prijať "hotové" z prostredia. Auxotrofné mutanty teda nie sú schopné rásť na minimálnych médiách, v ktorých tieto látky nie sú prítomné. Na druhej strane, keď prostredníctvom niektorého z týchto procesov dôjde k prijatiu DNA a jej využitiu, obnovuje sa u týchto buniek syntéza látok a bunky na minimálnom médiu vyrastú. Dochádza u nich k obnove prototrofie pre tieto esenciálne látky.
Genotyp auxotrofných mutantov sa tradične zapisuje skratkou príslušnej látky a znamienkom "-" (mínus), čo vyjadruje chýbanie, resp. mutáciu génu(ov) determinujúcich jej syntézu. Genotyp prototrofa potom prirodzene označujeme znamienkom "+" (plus). Príklad: Ak má kmeň E. coli genotyp trp- his+, bunky dokážu rásť na médiu bez prídavku histidínu ale nie bez prídavku tryptofánu. Ak je tryptofán pridaný do média, bunky tohto kmeňa vyrastú.
Na charakterizáciu auxotrofných mutantov používame série selektívnych médií, ktoré majú presne definované zloženie umožňujúce identifikovať určitý druh auxotrofie. Kompletné médiá obsahujú všetky nutričné zložky (aminokyseliny, dusíkaté bázy, vitamíny, atď.), na ktorých vyrastú aj auxotrofné mutanty. Používajú sa na premnoženie (zväčšenie biomasy) bakteriálnych kolónií.
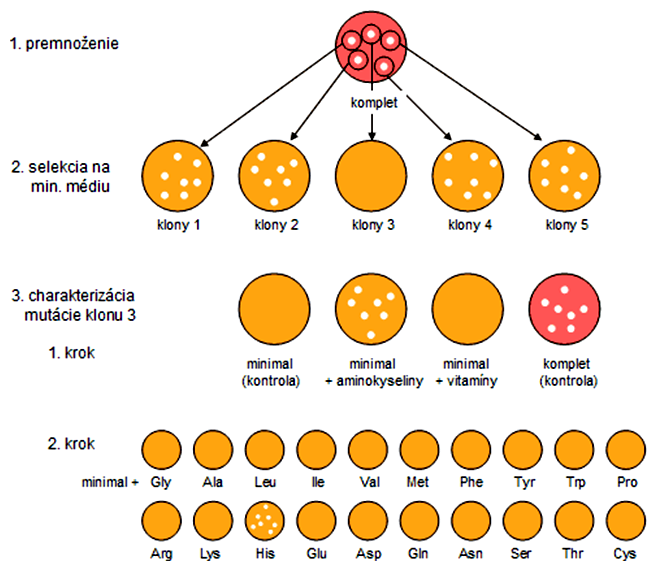
- premnoženie zmesi kolónií (rôznych genotypov) a odpichnutie jednotlivých kolónií na minimálne médiá
- identifikácia auxotrofných mutantov (klon 3)
- (krok 1) charakterizácia auxotrofie výsevom na minimálne médiá obohatené o určitú skupinu látok (aminokyseliny); kontrolné výsevy sa často používajú v experimentoch na vylúčenie pokusných chýb
- (krok 2) charakterizácia auxotrofie výsevom na minimálne médiá obohatené len o konkrétne aminokyseliny (histidín)
Konjugácia link
Konjugácia predstavuje jednosmerný prenos DNA z donorovej bunky do bunky recipientnej prostredníctvom špeciálnych štruktúr, tzv. konjugačných pilusov (sex-pilusov). Tento proces vyžaduje priamy bunkový kontakt, a z toho dôvodu nie je ovplyvnený prítomnosťou nukleáz v kultivačnom médiu. Recipientná bunka, ktorá prijala DNA fragment a zahrnula ho do svojho metabolizmu, sa nazýva transkonjugant. Konjugácia baktérií je analogická kríženiam eukaryotických organizmov a nazýva sa aj parasexuálny proces. Výmena génov nastáva rekombináciou, ktorá nie je reciproká ako u eukaryotov, ale prebieha len z plazmidu do bakteriálneho chromozómu pomocou jedného (ak je plazmid kruhová molekula) alebo dvoch crossing-overov (ak je donorová molekula lineárna). Zistilo sa, že za túto konjugáciu sú u kmeňa E. coli K12 zodpovedné konjugatívne plazmidy (napr. RK2 plazmid, F plazmid).
F faktor link
Konjugácie sú schopné len bunky, ktoré vlastnia tzv. F faktor (F pre fertilitu, čiže plodnosť). Je to plazmidová DNA, ktorá obsahuje informáciu pre syntézu spomenutých sex-pilusov a ďalších molekúl potrebných pre transfer DNA. F plazmid sa prenáša len z donorových buniek (označené F+) do recipientných, ktoré tento plazmid nemajú (F-). Pri prenose F plazmidu dochádza zároveň k jeho replikácii, takže z donorovej bunky sa F plazmid nevytráca. Následne po konjugácii získajú recipientné bunky fenotyp F+ a samy môžu byť donorom tohto plazmidu pre ďalšie bunky. Prenos F plazmidu trvá u E. coli približne 5 min.
Hfr kmene link
F plazmid existuje v bunkách ako samostatne sa replikujúca kruhová molekula DNA o veľkosti 95 kbp. V niektorých prípadoch môže dochádzať k jej včleneniu do bakteriálneho chromozómu na niekoľkých potenciálnych špecifických miestach. DNA, ktorá je schopná sa integrovať do hostiteľského chromozómu, sa nazýva epizóm. Takéto kmene označujeme Hfr (angl. high frequency of recombination).
Podobne ako F+ bunky tak aj Hfr bunky sú schopné iniciovať konjugáciu s tým rozdielom, že pri prenose DNA z donorovej bunky do recipientnej sa prenáša celý bakteriálny chromozóm, ktorého je F plazmid súčasťou. Pri konjugácii Hfr kmeňov nezískavajú recipientné bunky fenotyp F+. Je to tak z toho dôvodu, lebo gény pre syntézu konjugatívnych pilusov sa nachádzajú z pohľadu vedúceho vlákna prenášaného do recipientnej bunky bližšie k 3'-koncu F plazmidu. Keďže sa miesto, kde sa štiepi jednoreťazcová DNA plazmidu, nachádza bližšie k 5'-koncu, a plazmid je súčasťou bakteriálneho chromozómu, je potrebný transfer celého chromozómu, aby nastal aj prenos génov F plazmidu, ktoré sú pri jeho 3'-konci. Tento proces trvá približne 100 min, pričom je to dosť dlhá doba na to, aby sa v ktorejkoľvek fáze predčasne ukončil.
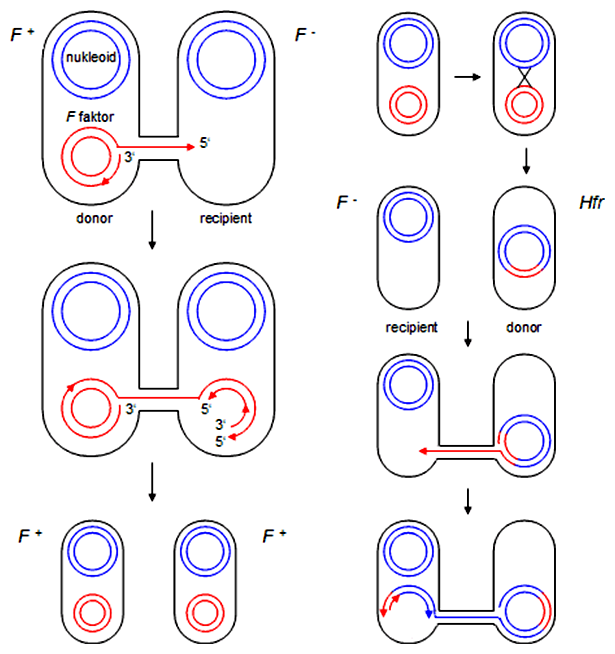
F' kmene link
F plazmid integrovaný v bakteriálnom chromozóme má možnosť sa z chromozómu znovu vyštiepiť. Môže sa však stať, že toto štiepenie prebehne nepresne a F plazmid si "odnesie" aj niekoľko génov v tesnej blízkosti miesta, do ktorého sa v bakteriálnom chromozóme integroval. V takomto prípade sa takýto plazmid označuje F' (čítaj 'ef s čiarou') , pričom sa pri jeho označení zvyknú zapisovať aj gény, ktoré si tento plazmid z chromozómu "ukradol". Napr. F'(lac) je F plazmid, ktorý sa nepresne vyštiepil z bakteriálneho chromozómu spolu s príslušným čítacím rámcom laktózového operónu. Konjugáciou F' donorových buniek s recipientnými F-, získajú bunky fenotyp F', ktorý je de facto typu F+ (schopné konjugácie). Prenos F' plazmidu do F- recipientných buniek označujeme aj pojmom sex-dukcia.
Prokaryotická bunka je haploidná, resp. monoploidná, pretože obsahuje len jeden bakteriálny chromozóm. Hfr a F' kmene sú vhodné na štúdium vzťahov dominancie a recesivity génov, pretože sa v bunke nachádzajú dve alely: jednak alela na bakteriálnom chromozóme a jednak na plazmide. Stav, kedy sú v prokaryotickej bunke prítomné dve alely (jedna na pôvodnom bakteriálnom chromozóme a druhá na vnášanej DNA do recipientnej bunky konjugáciou), označujeme pojmom merodiploid.
Transdukcia link
Transdukcia predstavuje prenos DNA medzi bunkami prostredníctvom bakteriálnych vírusov - bakteriofágov, alebo skrátene len fágov. Využitím nepresností v nabaľovaní DNA do vírusových častíc - viriónov - možno docieliť transfer vlastných génov bakteriálnej bunky do iných buniek práve prostredníctvom infekcie fágmi. S určitou pravdepodobnosťou sa totiž stáva, že fágy do svojich proteínových hlavičiek nenabalia vlastnú zreplikovanú DNA, ale cudziu, ktorú potom úspešne prenášajú do iných buniek ako vlastnú. Bunky, ktoré prijali od fágov cudzorodú DNA, sa nazývajú transduktanty. Keďže DNA je počas transdukcie chránená fágovou časticou, nie je tento proces citlivý k nukleázam. Transdukciu sprostredkúvajú fágy, takže k prenosu DNA nie je potrebný bunkový kontakt.
V princípe možno odlíšiť dve kategórie fágov. Lytické fágy sú agresívne fágy, ktoré po infekcii bakteriálnej bunky spôsobujú degradáciu bakteriálnej DNA a následnú deštrukciu (lýzu) buniek. Tieto fágy môžu nabaliť ľubovoľné časti bakteriálnej DNA, pokiaľ majú požadovanú veľkosť blízku veľkosti fágového genómu. Sem patria napr. T-párne a nepárne fágy (T1, T4, T7 fág). Spôsob, akou lytické fágy prenášajú medzi bunkami DNA, nazývame generalizovaná transdukcia.
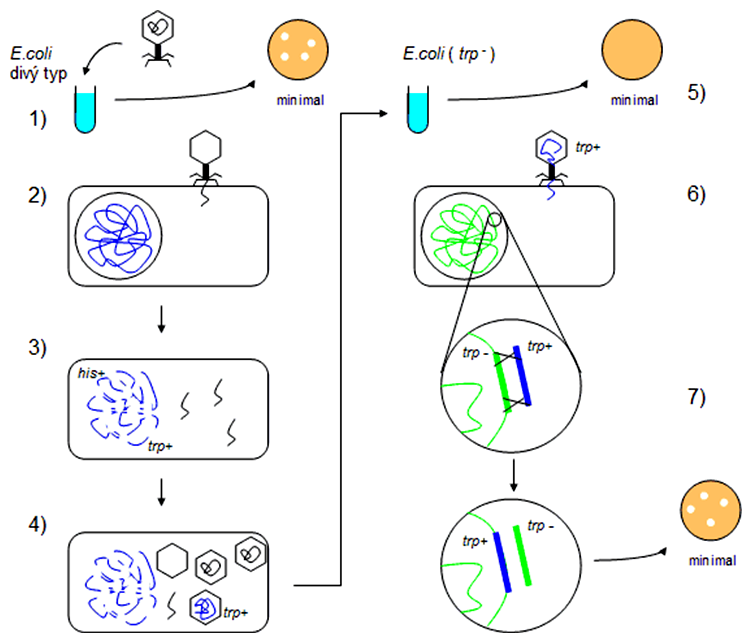
- kultúra divého typu E. coli, ktorá rastie na minimálnom médiu
- infekcia tejto kultúry fágom, transfer fágovej DNA do bunky
- replikácia fágovej DNA a degradácia hostiteľskej DNA
- tvorba fágových hlavičiek, do ktorých sa vbaľuje DNA (fágová aj hostiteľská), nasleduje lýza buniek
- kultúra kmeňa E.coli auxotrofného na tryptofán - nerastie na minimálnom médiu
- infekcia tohto kmeňa fágovým lyzátom z kroku 4.
- rekombinácia na tryptofánovom lokuse dvomi crossing-overmi a zámena fenotypu trp- za trp+ - bunky rastú na minimálnom médiu
Lyzogénne (temperované) fágy naopak využívajú stratégiu integrácie do bakteriálneho chromozómu (epizóm) na špecifických miestach, pričom množia sa súčasne s delením bakteriálnych buniek. Na určitý podnet potom indukujú lýzu buniek podobne ako lytické fágy. Rozdiel v transdukcii je ten, že takéto fágy môžu vbaliť do hlavičiek len takú cudziu DNA, ktorá sa nachádza v blízkosti miesta ich inzercie do bakteriálneho genómu (podobná situácia ako v prípade F' plazmidu pri konjugácii). Najpoužívanejším temperovaným fágom je fág λ (lambda). Lyzogénne fágy prenášajú DNA medzi bunkami spôsobom špecifickej transdukcie.
Transformácia link
Pri transformácii sa jedná o transfer holej DNA v médiu do bakteriálnych buniek bez konkrétnych vektorov (napr. fágy). Tento proces je citlivý voči nukleázam, pretože DNA nie je žiadnym spôsobom chránená. Bunkový kontakt pre transformáciu nie je potrebný. Bunky sa prijatiu cudzej DNA snažia za normálnych okolností vyhnúť, takže ich stena nie je pre DNA priepustná (DNA k tomu ani nemá chemické predpoklady). Okrem toho sa v bunke nachádza účinný restrikčno-modifikačný systém, ktorý pozostáva z enzýmov degradujúcich cudzorodú DNA a chrániacich vlastnú DNA pre degradáciou.
Za určitých okolností sa však môže stať, že bunky sa stávajú kompetentné, a teda ochotné prijať cudzorodú DNA. Každý druh baktérie požaduje iné podmienky pre vznik kompetencie. Bunky, ktoré úspešne prijali cudzorodú DNA pomocou transformácie, sa nazývajú transformanty. Bacillus subtilis sa stáva kompetentným pri prechode z exponenciálnej fázy do stacionárnej fázy, keď dochádzajú v prostredí živiny. Prijatie cudzorodej DNA je stratégia, akou by mohol získať výhodné gény na prekonanie nepriaznivého obdobia.
Naproti tomu Escherichia coli je značne imúnna voči prirodzenej transformácii. Na prekovanie tejto bariéry sa potom používajú techniky molekulárnej biológie, medzi ktoré patrí chemická transformácia a elektroporácia. V prvom prípade sa využitím dvojmocných katiónov (napr. CaCl2) a teplotných zmien indukujú v membráne póry a kanály, cez ktoré potom DNA preniká dovnútra bunky (okrem toho je nutné použiť bakteriálne kmene s defektným restrikčno-modifikačným systémom). Elektroporáciu zabezpečuje špeciálny prístroj - elektroporátor - ktorý permeabilizuje membránu pomocou silných elektrických impulzov. Kompetencia buniek sa tým rádovo zvyšuje.
Mapovanie génov prokaryot link
Ľubovoľný z týchto troch uvedených procesov možno využiť na mapovanie génov prokaryot, tzn. určovať ich pozíciu a vzdialenosti na DNA. Pri mapovaní génov pomocou konjugácie používame najčastejšie Hfr ako donorový kmeň. Recipientný kmeň (F-) má rôzne definované mutácie, aby bolo možné po konjugácii a následnej rekombinácii s "funkčnými" génmi donorového kmeňa selektovať transkonjugantov na príslušných selektívnych médiách. Princíp mapovania spočíva v rôznych časoch, v ktorých prerušujeme konjugáciu (postačuje k tomu silnejšie pretrepanie bunkovej suspenzie). Vzdialenosti medzi génmi sú potom dané časovými jednotkami, kedy transkonjuganty získali nový fenotyp od Hfr kmeňa.
Mapovanie génov pomocou generalizovanej transdukcie a transformácie je v princípe rovnaký. Môže sa nimi zisťovať predovšetkým sila väzby, tzn. relatívna vzdialenosť medzi dvoma génmi. Na selekciu transduktantov a transformantov používame recipientné kmene mutantné v dvoch (najjednoduchší prípad) analyzovaných génoch, medzi ktorými chceme zistiť vzdialenosť. Princíp mapovania spočíva v tom, že čím je vzdialenosť medzi génmi menšia, o to s väčšou frekvenciou vznikajú transduktanty a transformanty v obidvoch génoch súčasne, pretože sa prenášajú ako celok. Súčasný prenos dvoch alebo viacerých génoch v závislosti od použitého spôsobu mapovania označujeme pojmom kotransdukcia alebo kotransformácia.
Špecializovanú transdukciu možno len ťažko využiť na mapovanie génov, pretože pomocou lyzogénnych fágov možno preniesť len veľmi krátke kusy DNA, ktoré sa nachádzajú v okolí miesta inzercie fágu do donorovej DNA. Na druhej strasne však možno využiť túto ich špecifickú integráciu do definovaného miesta na chromozóme na transfer konkrétnych (teda nie náhodných) génov nachádzajúcich sa v blízkosti takejto sekvencie.