Autor: Peter Pančík
Publikované dňa:
Citácia: PANČÍK, Peter. 2016. Biopedia.sk: Populačná genetika. [cit. 2024-04-25]. Dostupné na internete: <https://biopedia.sk/genetika/populacna-genetika>.
V genetike sa pod pojmom populácia rozumie skupina jedincov toho istého druhu, ktoré obývajú presne vymedzený areál, v ktorom sa každý jedinec môže potenciálne párovať s ľubovoľným iným členom populácie (pozri upozornenie nižšie). Takáto populácia sa zvykne niekedy označovať ako mendelistická populácia. Jednou zo základných vlastností prírodných populácií je, že organizmy v nich žijúce sa odlišujú vo fenotype v mnohých znakoch. Úlohou populačnej genetiky je určiť mieru tejto genetickej premenlivosti v rámci populácie a medzi populáciami, vysvetliť jej pôvod a popísať faktory, ktoré ju v priebehu evolúcie ovplyvňujú.
Populačná genetika určuje genetickú variabilitu prostredníctvom určenia genetickej štruktúry populácie. Tá spočíva v meraní frekvencie alel konkrétnych génov, ktoré tento rozdielny fenotyp determinujú. Alelické frekvencie teda predstavujú percentuálne zastúpenie určitej alely génu v populácii. Od alelických frekvencií treba odlišovať genotypové frekvencie, ktoré predstavujú percentuálne zastúpenie nie jednotlivých alel, ale konkrétneho genotypu v populácii. Príklad: Ak sa v populácii vyskytuje alela pre modré oči a a hnedé oči A, ktoré sú rozdielnymi formami toho istého génu, alelické frekvencie predstavujú podiel výskytu alely a (alebo A) v populácii a genotypové frekvencie predstavujú zastúpenie genotypov aa, Aa a AA v populácii.
Hardyho-Weinbergov zákon link
Pre potreby populačnej genetiky bolo potrebné zaviesť zjednodušený matematický model, ktorý by umožnil kvantitatívne vyjadrenie genetickej variability. Jedným z nich je Hardyho-Weinbergov zákon (ďalej H-W zákon), ktorý vyjadruje vzťah medzi alelickými a genotypovými frekvenciami v populácii. Tento zákon sformulovali v roku 1908 anglický matematik GODFREY HAROLD HARDY (1877-1947) a nemecký fyziológ WILHELM WEINBERG (1862-1937).
Použitie H-W zákona pre konkrétnu populáciu vyžaduje splnenie niekoľkých podmienok, z ktorých som už niektoré spomenul:
- párovanie jedincov v populácii je náhodné
- druh sa rozmnožuje pohlavným spôsobom
- analyzovaný organizmus by mal byť diploidný
- uvažovaný gén má dve alely
- alelické frekvencie sú rovnaké u samčieho aj samičieho pohlavia
- veľkosť populácie je veľmi veľká (500 a viac jedincov)
- generácie nasledujú v rade za sebou
- alelické frekvencie sa nemenia vplyvom evolučných síl (mutácie, migrácia, selekcia)
Ak sú tieto podmienky splnené, platí H-W zákon a populácia sa nachádza v tzv. H-W rovnováhe, pretože sa v nej nemenia alelické frekvencie.
Systém kríženia link
Pre využitie H-W zákona je dôležité, aby mohlo dochádzať k ľubovoľnej kombinácii genotypov, tzn. ako už bolo povedané, aby sa každý jedinec mohol potenciálne párovať s hociktorým iným jedincom. Tým sa zabezpečí, že nedôjde k preferovaniu určitého genotypu, čím by sa v konečnom dôsledku po mnoho generácii menili alelické a genotypové frekvencie. Taký systém párovania, ktorý umožní H-W rovnováhu, sa nazýva náhodné párovanie, čiže panmixia. Panmiktická populácia je mendelistická populácia, v ktorej sa dva genotypy párujú s takou frekvenciou (pravdepodobnosťou), v akej frekvencii sa oba genotypy nachádzajú v populácii. Príklad: Dané sú genotypové frekvencie AA 25%, Aa 50% a aa 25%. Pravdepodobnosť, že AA sa spáruje s Aa je 0,25 × 0,5 = 0,125 (12,5%) a pod.
2. Ak počítame alelické a genotypové frekvencie v panmiktickej populácii, vždy sa zameriavame na určité konkrétne gény a genotypy (napr. znak pre farbu očí v predchádzajúcom texte). Náhodné párovanie neprebieha vo všetkých znakoch naraz, pretože tak by žiadna populácia nebola panmiktická (výška, národnosť, farba pleti u ľudí)!
Ostatné systémy kríženia predstavujú problém pre H-W zákon. Inbríding (angl. inbreeding) predstavuje kríženie medzi príbuznými (príbuzenské kríženie). V ľudských spoločenstvách k nemu dochádza zvyčajne na stupni bratranec - sesternica (prvostupňoví). Pri mnohých rastlinných druhoch dochádza pravidelne k samoopeleniu rovnako aj u hmyzu dochádza ku kríženiu brat - sestra. Indríding však nevyhnutne nevyžaduje spojenie blízkych príbuzných. V málo početných populáciách sa tiež vyskytuje istý stupeň inbrídingu, pretože ich členovia zdieľajú minulých alebo súčasných predkov. Homogamia (angl. assortative mating) je príkladom nenáhodného párovania, pri ktorom dochádza k preferenčnému vyhľadávaniu partnera s podobným genotypom (IQ, výška). Obidva tieto spôsoby párovania menia genotypové frekvencie, ale nemenia alelické frekvencie!
Alelické frekvencie link
Ak dominantnú alelu označíme písmenom A a recesívnu alelu a, potom frekvencia alely A v populácii je p a frekvencia alely a je q, pričom platí H-W rovnováha:
p(A) + q(a) = 1
Alelické frekvencie môžeme vypočítať priamo z absolútnych počtov genotypov, pričom si treba uvedomiť: 1) dominantný homozygot má dve alely A a heterozygot len jednu alelu, 2) jedná sa o diploidné organizmy, takže každý jedinec má dve alely:
| p(A) = | 2 × počet AA + počet Aa |
| 2 × počet všetkých jedincov |
Genotypové frekvencie link
Genotypové frekvencie dostaneme rozvinutím binómu:
(p + q)2 = 1
teda
p2(AA) + 2pq(Aa) + q2(aa) = 1
Ak poznáme absolútne počty genotypov, jednotlivé genotypové frekvencie predstavujú podiel počtu jedincov konkrétneho genotypu k súčtu všetkých jedincov, pričom súčet všetkých frekvencií je rovný 1:
| p2(AA) = | počet AA |
| počet všetkých jedincov |
Distribúcia alel v populácii link
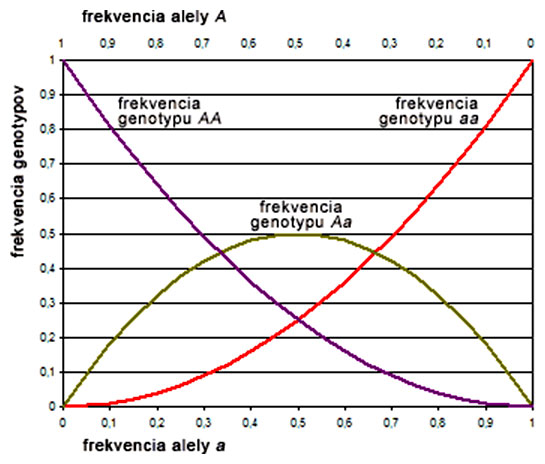
Nasledovný graf vyjadruje vzťah medzi alelickými a genotypovými frekvenciami v populácii, v ktorej platí H-W zákon. Jednotlivé hodnoty genotypových frekvencií pre konkrétnu populáciu sa nachádzajú v priesečníkoch farebných kriviek s kolmicou spustenou z niektorého bodu na osi x, ktorý predstavuje hodnotu alelických frekvencií. Všimnite si, že ak platí H-W rovnováha, frekvencia heterozygotov nie je nikdy vyššia ako 50%.
Testovanie H-W rovnováhy link
V tomto prípade chceme zistiť, či nejaká populácia, resp. nami vybraná vzorka z tejto populácie, spĺňa podmienky H–W rovnováhy. To predpokladá, že na ňu nepôsobia vonkajšie faktory, ktoré ovplyvňujú alelické a genotypové frekvencie (hlavne selekcia a genetický drift). Nasledovný vzorec na výpočet populačnej rovnováhy môžeme použiť len v prípade, ak pre sledovaný alelový pár je typický intermediárny alebo kodominantný typ dedičnosti, a teda presne vieme, akú časť populácie tvoria dominantní homozygoti, heterozygoti a recesívni homozygoti:
2pq(Aa) = 2 × √[p2(AA) × q2(aa)]
Pri dôkladnom testovaní H–W rovnováhy treba v prvom rade vypočítať alelické frekvencie v populácii z absolútnych počtov jednotlivých genotypov, ako už bolo uvedené, a potom zvoliť vhodnú nulovú hypotézu, ktorú otestujeme pomocou Χ2–testu:
| p(A) = | 2 × počet AA + počet Aa |
| 2 × počet všetkých jedincov |
Multialelické lokusy link
H-W zákon rieši aj stav, kedy sa v populácii môže na jednom lokuse vyskytovať viacero alel (multialelické lokusy). Spravidla sa označujú spoločným písmenom s číselným indexom. Ak sú na jednom lokuse prítomné tri alely A1, A2, A3, platí:
p(A1) + q(A2) + r(A3) = 1
Genotypové frekvencie vypočítame obdobne ako v prípade dvoch alel:
[p(A1) + q(A2) + r(A3)]2 = p2(A1A1) + 2pq(A1A2) + q2(A2A2) + 2pr(A1A3) + 2qr(A2A3) + r2(A3A3) = 1
X-viazané gény link
Výpočet alelických a genotypových frekvencií lokusov viazaných na chromozóm X je komplikovanejší o fakt, že samce majú len jeden chromozóm X, zatiaľčo samice ich majú dva (cicavce, drozofila). Napriek tomu princíp výpočtu je rovnaký. U samíc je výpočet oboch frekvencií ekvivalentný s autozómovo viazanými lokusmi. U samcov, keďže sú hemizygotné (t.j. majú len jeden chromozóm z chromozómového páru namiesto štandardných dvoch), sa alelické frekvencie rovnajú genotypovým frekvenciám. Pre populácie samíc a samcov platí nasledovná H-W rovnováha:
Populácia ♀
alelické frekvencie: p(XA) + q(Xa) = 1
genotypové frekvencie: p2(XAXA) + 2pq(XAXa) + q2(XaXa) = 1
Populácia ♂
alelické frekvencie: p(XA) + q(Xa) = 1
genotypové frekvencie: p(XA) + q(Xa) = 1
Alelické frekvencie X-viazaných génov v zmiešanej populácii (samice + samce) možno taktiež vypočítať z absolútnych počtov genotypov, treba však rozlišovať, či sa jedná o samice alebo samce:
| p(XA) = | 2 × počet XAXA samíc + počet XAXa samíc + počet XAY samcov |
| 2 × počet všetkých samíc + počet všetkých samcov |
Genotypové frekvencie sa počítajú pre každé pohlavie zvlášť, tak ako to uvádzajú dva odlišné vzorce H-W rovnováhy v horeuvedenej tabuľke.
Dôležitým poznatkom vyplývajúcim z voľnej kombinovateľnosti X-viazaných génov je, že fenotypy, ktoré sú podmienené recesívnou alelou budú oveľa početnejšie pri mužskom ako pri ženskom pohlaví. Rozdiely medzi početnosťou recesívnych genotypov pri ženskom a mužskom pohlaví sa budú tým zväčšovať, čím sa recesívna alela stáva zriedkavejšou.
Odchýlky od H-W rovnováhy link
Porušenie H-W rovnováhy je dôsledkom pôsobenia faktorov, ktoré menia alelické a genotypové frekvencie v populácii. Okrem už spomínaných systémov kríženia (inbríding, homogamia) sú hlavnými a dôležitejšími dve skupiny činiteľov:
- systematické činitele
- mutácie
- migrácia
- selekcia
- stochastické činitele
- genetický drift
Mutácie link
Mutácie sú najdôležitejším evolučným činiteľom, pretože len pomocou nich vznikajú nové alely, ktoré sa prostredníctvom ostatných činiteľov v populácii rozšíria, alebo naopak zaniknú. Mutácie sú prevažne recesívne, pretože väčšina ich predstavuje nevýhodu pre organizmus. Tie, ktoré sú pre organizmus výhodné, sú v populácii udržané a rozširované do ďalších generácií. Výhoda, nevýhoda či neutralita mutácií závisí od konkrétneho prostredia, v ktorom organizmus žije. Väčšina alel mutuje vo veľmi nízkej frekvencii, ktorá sa pohybuje v rozsahu 10-4 až 10-6 nových mutácií na gén za generáciu, čo je zhruba 1 mutácia na 100 000 alel za generáciu. Čím väčšia je populácia, tým väčšia je pravdepodobnosť vzniku novej mutácie.
Od priamych mutácií, ktoré menia štandardnú alelu na mutantnú, treba odlišovať spätné mutácie, ktoré menia mutantnú alelu späť na štandardnú. Z toho logicky vyplýva, že frekvencia spätných mutácií je nižšia, pretože zatiaľčo priame mutácie si "môžu vybrať" gén, ktorý zmutuje, spätné mutácie sa "musia zacieliť" na jeden konkrétny gén a zmeniť ho na pôvodný. Mutácie si bunka nevyberá, nie sú teda cieleným nástrojom, ktorý si organizmus vytvára, aby sa adaptoval na nové podmienky v prostredí. Jediné čo organizmus môže, je uchovať si náhodnú žiadúcu mutáciu.
Ak si pravdepodobnosť, že alela A zmutuje na a označíme μ, a spätnú mutáciu (a na A) označíme ν, potom zmena alelickej frekvencie Δp po prvej generácii je daná vzťahom:
Δp = νq - μp
Príklad: Ak frekvencia priamej mutácie je 5.10-5, frekvencia spätnej mutácie je 2.10-5, alelické frekvencie p = 0,9 a q = 0,1, potom zmena alelickej frekvencie Δp je:
Δp = (2.10-5 × 0,1) - (5.10-5 × 0,9) = -0,000043
Vidíme, že mutácie samy o sebe menia alelické frekvencie len nepatrne.
Migrácia link
Migráciu označujeme aj pojmom génový tok (angl. gene flow). Ak dochádza k premiestňovaniu jedincov medzi populáciami, nastávajú zmeny alelických a genotypových frekvencií. Efekt migrácie na zmenu genetickej variability ovplyvňuje veľkosť migrovanej skupiny a rozdiel v alelických frekvenciách medzi populáciami. Výsledkom môže byť aj prínos novej alely, ktorá v druhej populácii chýba. Rozdiely medzi oboma skupinami sa po niekoľkých generáciách náhodného kríženia zmenšujú.
Ak si frekvenciu alely A v migrujúcej skupine x označíme px, frekvenciu tej istej alely v donorovej populácii y označíme py a percento imigrantov, ktoré tvorí určitý podiel z recipientnej skupiny, ako m, potom zmena alelickej frekvencie Δp po migrácii bude:
Δp = m(px - py)
Príklad: Alelická frekvencia p v migrujúcej skupine, ktorá pozostáva z 10% jedincov pôvodnej recipientnej skupiny, je 0,5, alelická frekvencia p v donorovej skupine je 0,8.
Δp = 0,1 × (0,5 - 0,8) = -0,03
Selekcia link
Prírodná selekcia sa niekedy označuje pojmom Darwinovská selekcia, pretože je základom evolučnej teórie, ktorej autorom je CHARLES DARWIN (1809-1882). Darwin vo svojom diele O pôvode druhov cestou prírodného výberu (1858) popísal fungovanie selekcie na prírodné populácie, pričom použil termíny ako "boj o život" a "prežívanie najlepšie prispôsobených" (survival of the fittiest). Podľa jeho koncepcie je nielen dôležité, aby organizmus vytvoril mnoho potomkov, ale aby sa zároveň čo najviac týchto potomkov dožilo reprodukčného veku. Čiže plodnosť je rovnako dôležitá ako prežívanie. Taktiež zdôraznil, že prírodná selekcia pôsobí na jednotlivca a nie na druh.

Väčšina druhov produkuje viac potomkov, ako je schopných prežiť do reprodukčného veku, pričom je to do veľkej miery dané ich genetickou výbavou. Prírodná selekcia uprednostňuje jedincov s takým genotypom, ktorý najlepšie obstojí v daných podmienkach prostredia, takže sa také genotypy prenášajú do ďalších generácií a v populácii sa rozširujú. Selekciou sa tak menia alelické aj genotypové frekvencie. Postupné genetické zlepšovanie populácie, ktoré je výsledkom prírodnej selekcie je základom procesu, ktorý sa označuje ako evolučná adaptácia.
Prírodná selekcia sa meria na základe relatívnej reprodukčnej schopnosti genotypu, ktorá sa nazýva fitness alebo viabilita a označuje sa W. Genotyp, ktorý zabezpečí najlepšie prežívanie a reprodukciu svojich potomkov má fitness 1, pričom relatívny fitness ostatných genotypov sa určí porovnaním s týmto genotypom. Selekčný koeficient (s) je naproti tomu relatívna miera, akou prírodná selekcia pôsobí proti genotypu, pričom platí:
s = 1 - W
| G1G1 | G1G2 | G2G2 | |
| počet jedincov v reproduktívnom veku | 16 | 10 | 20 |
| počet potomkov v ďalšej generácii | 128 | 40 | 40 |
| priemerný počet potomkov na jedného rodiča | 128 / 16 = 8 | 40 / 10 = 4 | 40 / 20 = 2 |
| fitness | 8 / 8 = 1 | 4 / 8 = 0,5 | 2 / 8 = 0,25 |
| selekčný koeficient | 1 - 1 = 0 | 1 - 0,5 = 0,5 | 1 - 0,25 = 0,75 |
Selekcia môže uprednostňovať ľubovoľný genotyp, od čo bude závisieť aj aký bude mať efekt na alelické a genotypové frekvencie a na populačnú rovnováhu. Účinok selekcie závisí aj od počiatočnej hodnoty alelickej frekvencie. V nasledovnej tabuľke sa pokúsim zhrnúť všetky možné varianty, ako môže selekcia pôsobiť na genotypy (s je selekčný koeficient genotypu AA, t je selekčný koeficient genotypu aa):
| typ selekcie | fitness genotypov | zmena alelickej frekvencie | |||
|---|---|---|---|---|---|
| bez selekcie | W11 = W12 = W22 = 1 | Δp = Δq = 0 | |||
| proti recesívnemu homozygotovi | W11 = W12 = 1; W22 < 1 |
|
|||
| proti dominantnej alele | W11 = W12 < 1; W22 = 1 |
|
|||
| bez dominancie | W11 < W12 < 1; W22 = 1 |
|
|||
| favorizujúca heterozygotov | W11 a W22 < 1; W12 = 1 |
|
|||
| proti heterozygotom | W12 < (W11 = W22 = 1) |
|
Prípady, kedy selekcia vedie k eliminácii alebo aspoň značnej redukcii frekvencie niektorej alely, nazývame usmernenou selekciou. Heterozygotná nadradenosť alebo superdominancia je špeciálnym prípadom selekcie, ktorá vedie k stabilizácii alelických frekvencií, pretože sa v populácii udržiavajú obidve alely. Príkladom zo života je kosáčikovitá anémia u ľudí. V homozygotne recesívnom stave spôsobuje smrť, pretože červené krvinky nie sú schopné normálne distribuovať kyslík do tkanív. Heterozygot sa prejavuje anémiou, ktorá vyvoláva komplikácie, ale človek prežije. Avšak v krajinách, ktoré sú postihnuté maláriou je kosáčikovitá anémia výhodná, pretože zabezpečuje imunitu voči pôvodcovi malárie (jednobunkový parazit množiaci sa vo vnútri červených krviniek). Homozygoti, ktorým chýba alela pre kosáčikovitú anémiu, podliehajú malárii.
Genetický drift link
Genetický drift je náhodný proces, ktorý ovplyvňuje alelické a genotypové frekvencie v malých populáciách. Princíp genetického driftu spočíva v tom, že z populácie vyberieme malú skupinu jedincov, ktorá sa potom rozmnožuje izolovane od pôvodnej populácie. Ku zmene alelických frekvencií dochádza preto, že vplyvom náhody sa do tejto skupiny dostane viac jedincov s určitou alelou ako je priemer pôvodnej populácie. Patrí sem tzv. efekt zakladateľa (angl. founder effect), keď malá skupina ľudí založí novú kolóniu, alebo bottleneck effect, keď dôjde k redukcii organizmov do takej miery, že ostane len niekoľko jedincov, ktoré sa znovu rozmnožia. Efekt genetického driftu nemožno presne vypočítať (keďže sa jedná o náhodný jav), možno však určiť aspoň interval, v akom dôjde k zmene alelických frekvencií. Dôležitú úlohu tu pritom zohráva efektívna veľkosť populácie, ktorá určuje počet jedincov v reprodukčnom veku, pričom závisí od pomeru samčích a samičích gamét.
| typ činiteľa | ovplyvnenie alelických frekvencií | genetická variácia medzi populáciami | genetická variácia v rámci populácie |
| inbríding | nemení | nemení | znižuje |
| mutácie | menia | môžu zvyšovať | zvyšujú |
| migrácia | mení | znižuje | zvyšuje |
| selekcia | mení | zvyšuje | mení |
| genetický drift | mení | môže meniť | znižuje |