Autor: Peter Pančík
Publikované dňa:
Citácia: PANČÍK, Peter. 2016. Biopedia.sk: Väzba génov. [cit. 2024-04-18]. Dostupné na internete: <https://biopedia.sk/genetika/vazba-genov>.
Dva gény, ktoré sa nachádzajú na nehomologických chromozómoch, majú rovnakú šancu dostať sa do ďalšej generácie prostredníctvom gamét nezávisle na sebe. Hovoríme u nich o voľnej kombinovateľnosti, ktorá je predpokladom vzniku mendelistických štiepnych pomerov. Ak sú však gény lokalizované na tom istom chromozóme, majú tendenciu dediť sa ako celok, a to tým viac, čím sa ich lokusy nachádzajú bližšie pri sebe. O takýchto génoch hovoríme, že sú vo väzbe a tvoria väzbovú skupinu.
Väzbu génov intenzívne študoval britský zoológ THOMAS HUNT MORGAN (1866-1945). Pri svojej práci s krížením drozofíl rozšíril poznanie dovtedajšej klasickej mendelistickej genetiky o princípy génovej väzby, ktoré sú známe ako Morganove pravidlá:
- gény lokalizované na jednom chromozóme sa dedia spoločne
- gény sú v chromozóme uložené lineárne
- medzi homologickými chromozómami môže prebehnúť rekombinácia

Rekombinácia link
Rekombinácia (crossing-over) je jav, ktorým sa vymieňajú úseky homologických chromozómov počas pachytene v profáze I. meiotického delenia pri vzniku pohlavných buniek. Miesta, kde dôjde k zlomu a výmene nesesterských chromatíd homologických chromozómov, sa nazývajú chiazmy. Frekvencia rekombinácie závisí od vzdialenosti génov na chromozóme.
Prostredníctvom rekombinácie sa vytvárajú nové typy gamét. Potomkovia, ktorí vykazujú rovnaký genotyp ako ich rodičia, sa volajú parentálne (rodičovské) genotypy. Ak dôjde počas tvorby gaméty k rekombinácii, vznikajú rekombinantné genotypy.
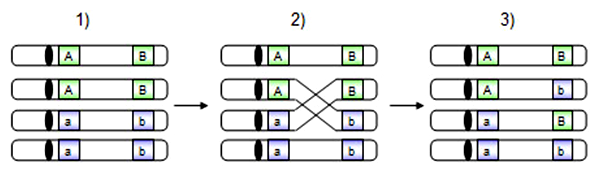
- Bunka v S-fáze obsahuje 2 súbory homologických chromozómov (modré a zelené), z ktorých každý má dve sesterské chromatidy. Bunka je genotypu AaBb.
- V profáze I. meiotického delenia dochádza ku konjugácii a rekombinácii medzi dvoma nesesterskými chromatidami homologických chromozómov.
- Po výmene úsekov vznikajú 2 parentálne gaméty s genotypom AB a ab a 2 rekombinantné gaméty s genotypom Ab a aB.
(čierna elipsa predstavuje centroméru, chromozómy so zelenými /resp. modrými/ alelami sú zreplikované sesterské chromatidy, modré a zelené chromozómy sú si navzájom homologické)
Podľa umiestnenia génov na chromozóme rozlišujeme tieto 3 prípady:
- gény nie sú vo väzbe - gény sú v takej vzdialenosti od seba, že rekombinantné gaméty vznikajú s rovnakou pravdepodobnosťou ako gaméty parentálneho typu
- neúplná väzba - vzdialenosť medzi génmi umožňuje, aby došlo k vzájomnej výmene nesesterských chromatíd a vzniku rekombinantných gamét
- úplná väzba - rodič vytvára len jeden typ gamét, pretože sú gény veľmi blízko seba, čím sa znižuje pravdepodobnosť vzniku crossing-overu medzi nimi
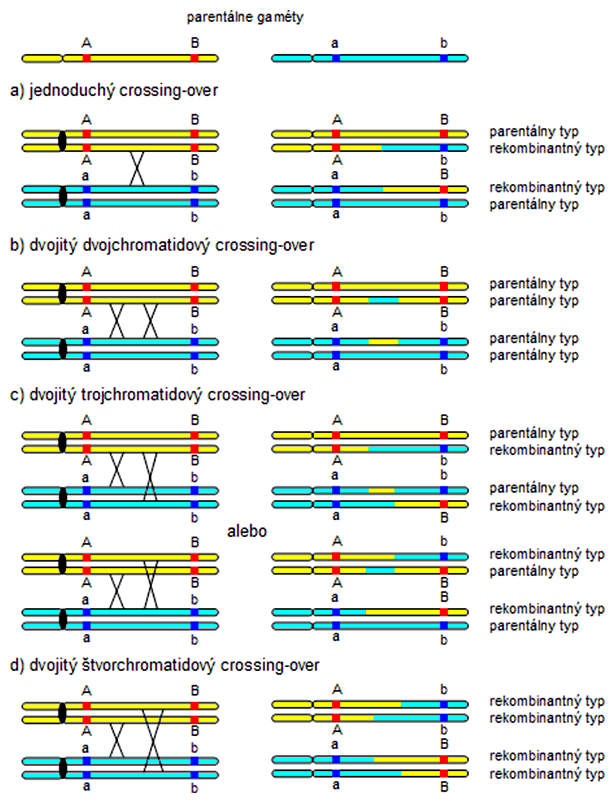
Dvojitý crossing-over link
Jednoduchý crossing-over medzi dvoma nesesterskými chromatidami homologických chromozómov produkuje najviac polovicu rekombinantných a polovicu parentálnych genotypov. Pri tvorbe genetických máp treba brať ohľad ako na jednoduchý tak aj na dvojitý crossing-over, keď dochádza k výmene úsekov medzi homologickými chromozómami na viacerých miestach.
Dvojitý crossing-over môže zahŕňať dve, tri alebo všetky štyri chromatidy. Ak prebehne medzi dvoma rovnakými nesesterskými chromatidami, výsledkom sú všetky štyri genotypy parentálneho typu. Ak dvojitý crossing-over zahŕňa tri chromatidy, vzniká 50% parentálnych a 50% rekombinantných genotypov. Ak rekombinácia zasiahne všetky štyri chromatidy, výsledkom je 100% rekombinantných genotypov. Ak vyhodnotíme veľké množstvo meióz na dvojitý crossing-over, výsledkom bude 50% parentálnych a 50% rekombinantných genotypov, čo dokazuje, že maximálny limit rekombinačnej frekvencie je 50% (ako pri voľnej kombinovateľnosti génov na rôznych nehomologických chromozómoch).
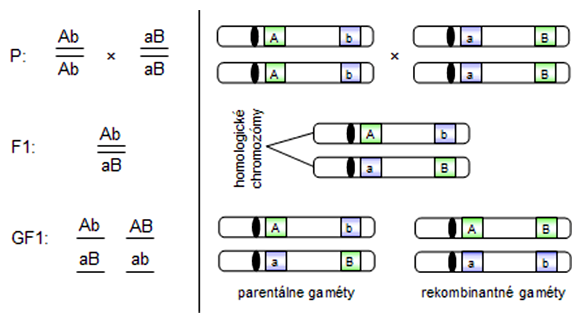
Symbolika link
Genotyp, ktorý je tvorený génmi vo väzbe, sa nezapisuje do riadku ale pod seba, pričom sú alely oddelené dvojitou čiarou naznačujúcou chromozóm. V prípade dvojitého heterozygota môžu nastať dva prípady usporiadania alel:
- cis-fáza (coupling) - dve dominantné alely sa nachádzajú na jednom chromozóme a dve recesívne alely na druhom
- trans-fáza (repulsion) - každý chromozóm nesie po jednej dominantnej a recesívnej alele

Mapovacie jednotky link
Na genetických mapách sa vzdialenosti nepočítajú tak ako sme zvyknutí z normálneho sveta fyzikálnych dĺžkových jednotiek. Vychádzajú totižto z genetických javov, predovšetkým crossing-overu, takže sa niekedy nazývajú aj väzbové mapy. Na kvantitatívne stanovenie vzdialenosti medzi dvoma génmi pri tvorbe väzbových (genetických) máp sa používa frekvencia, s akou vznikajú rekombinantné gaméty v porovnaní s rodičovskými. Úplnej väzbe zodpovedá frekvencia 0% (rekombinantné gaméty nevznikajú), voľnej kombinovateľnosti 50%. Jedna mapovacia jednotka zodpovedá 1% výskytu crossing-overu medzi dvoma génmi a nazýva sa centimorgan [cM] podľa objaviteľa väzby génov Morgana. Jeden centimorgan teda znamená takú vzdialenosť medzi dvoma génmi, ktorá dovolila vzniku crossing-overu a rekombinanta v 1 prípade zo 100.
Štiepne pomery pri väzbe vlôh sú iné ako pri voľnej kombinovateľnosti. Pri génovej väzbe nie sú štiepne pomery stabilné, ale sú podmienené silou väzby, ktorá je priamo úmerná vzdialenosti génov na chromozóme. Čím bližšie budú dva lokusy umiestnené voči sebe na chromozóme, tým silnejšia bude väzba, pretože pravdepodobnosť vzniku crossing-overu práve v mieste medzi týmito dvoma génmi je relatívne nízka.
Dvojbodový test link
V dvojbodovom testovacom krížení sledujeme vzdialenosť medzi dvoma génmi meraním frekvencie vzniku parentálnych a rekombinantných genotypov. Pri tomto teste dostaneme dva parentálne typy v približne rovnakých frekvenciách a dva rekombinantné typy, ktoré sa vyskytujú tiež približne v rovnakých frekvenciách. Vlastný fenotyp bude potom závisieť od usporiadania týchto dvoch alelických párov, t.j. či sú vo fáze cis alebo trans (pozri obrázky vyššie).
Na stanovenie sily väzby sa používa nasledovný vzorec:
| počet rekombinantov | × 100 = frekvencia rekombinantov [cM] |
| počet všetkých jedincov |
Z matematického hľadiska teda nejde o nič iné ako o určenie, koľko percent zo všetkých potomkov je po krížení rekombinantných. Dvojbodový test možno použiť pri mapovaní génov vtedy, ak gény ležia blízko pri sebe, pričom nezáleží na orientácii (ktorý z nich je bližšie k centromére) ale len na vzdialenosti medzi nimi.
Trojbodový test link
Hoci sa genetické mapy dajú robiť pomocou sérií dvojbodových testov, genetici spravidla mapujú viac génov naraz pomocou komplexnejších metód. Jednou z nich je trojbodový test pre tri gény, ktorý spočíva v krížení trojnásobného heterozygota s trojnásobným recesívnym homozygotom:
| ABC | × | abc |
| abc | abc |
Pri hodnotení testu je dôležité určiť v prvom rade poradie génov a ich väzbovú fázu (cis alebo trans). Ak mapujeme tri gény A, B a C, môže byť ich poradie na chromozóme ABC, ale aj ACB. Pri určení poradia vychádzame z frekvencie dvojitých rekombinantov, ktorých je najmenej, a fenotypu, ktorý takýto genotyp podmieňuje. Najpočetnejšie budú parentálne typy, u ktorých neprebehol crossing-over.
Vzdialenosť medzi génmi určíme taktiež pomocou frekvencie rekombinantov, pričom vychádzame z predpokladu, že výskyt crossing-overu je náhodný proces, a tak čím bude frekvencia rekombinantov nižšia, tým budú gény bližšie pri sebe.