Autor: Peter Pančík
Publikované dňa:
Citácia: PANČÍK, Peter. 2016. Biopedia.sk: Separácia proteínov - afinitná chromatografia. [cit. 2024-04-25]. Dostupné na internete: <https://biopedia.sk/molekularna-biologia/afinitna-chromatografia>.
Princípom afinitnej chromatografie je špecifická väzba (afinita) určitého štruktúrneho proteínového motívu so špecifickým ligandom, kovalentne naviazaným na matricu. Je to teda princíp väzby protilátok s antigénom - "kľúč a zámok". Proteíny bez tohto motívu sa za ideálnych podmienok na matricu neviažu, takže selektivita a vysoká rozlišovacia schopnosť, ako aj kapacita kolóny, sú pri tejto separačnej technike veľmi vysoké. V praxi je však často potrebné hľadať optimálne zloženie premývacieho roztoku (najmä s ohľadom na iónovú silu), ktorý odstráni nešpecificky naviazané proteíny. Väzba proteínu s matricou je dočasná (reverzibilná), takže so zmenou zloženia mobilnej fázy je proteín z kolóny vytesnený. Elučný roztok pôsobí nešpecificky, t.j. zmenou iónovej sily, pH alebo polarity, alebo vyviazanie proteínu spôsobuje väzba s kompetitívnym ligandom.
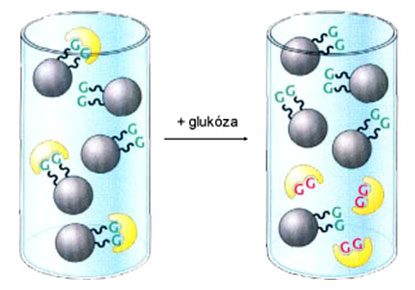
Príklad väzby proteínu na matricu s kovalentne naviazanou glukózou. Glukóza predstavuje pre daný proteín prirodzený substrát, takže tento fakt môže byť využitý pri jeho selektívnej purifikácii. Po odmytí ostatných kontaminujúcich proteínov možno predmetný proteín z matrice vytesniť (eluovať) roztokom glukózy.
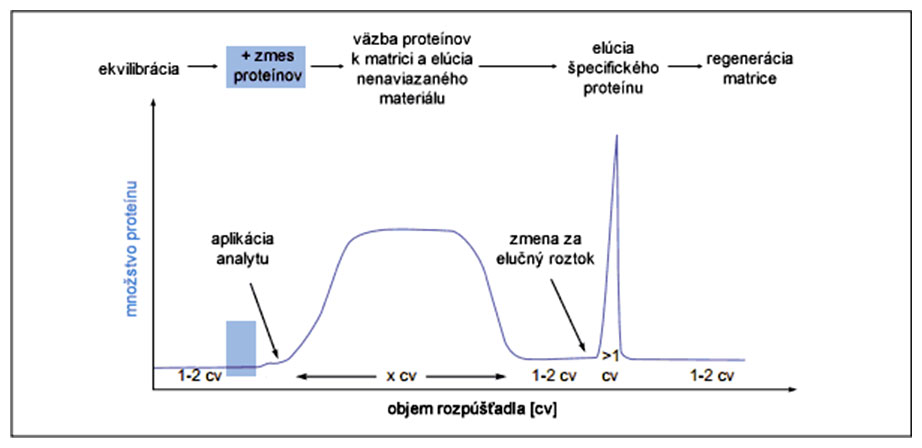
Po ekvilibrácii matrice a aplikácii analytu na kolónu dochádza k väzbe rekombinantného proteínu na matricu, pričom zvyšné proteíny kolónou len pretečú. (Následne možno pomocou premývacieho roztoku vytesniť z matrice aj nešpecificky naviazané proteíny.) Elúciu špecificky naviazaného proteínu spôsobí zámena tlmivého roztoku za elučný roztok. Matricu je zväčša potrebné po separácii regenerovať použitím špeciálneho roztoku.
V afinitnej chromatografii využívame niekoľko typov ligandov, ktoré možno použiť na purifikáciu špecifických proteínov alebo skupín proteínov. Vo všeobecnosti sa jedná o purifikáciu proteínov na základe ich reverzibilnej väzby s ligandom, ktorým je buď prirodzený substrát alebo jeho derivát (analóg).
| ligand matrice | afinita k | poznámka |
| imunoglobulíny | špecifický substrát | Imunoafinitná chromatografia. IgG sú veľmi často viazané na matricu prostredníctvom väzby s rekombinantným bakteriálnym proteínom A alebo G, ktoré špecificky viažu Fc reťazce IgG rôzneho pôvodu. Elúcia proteínu (antigénu) z protilátky pomocou nízkeho pH (<2,7). Antigénom môžu byť aj väčšie častice, napr. virióny alebo bakteriálne bunky. |
| lektíny (napr. konkanavalín A) | glykoproteíny | Lektíny sú proteíny, ktoré viažu niektoré monosacharidy. Štúdium glykozylácie proteínov. |
| nukleová kyselina | proteíny viažúce sa na DNA | Vhodné napr. na purifikáciu transkripčných faktorov. |
Purifikácia rekombinantných proteínov - proteínové kotvy link
Nie každý proteín je možné prirodzene separovať pomocou tejto techniky. Využitie afinitnej chromatografie je predovšetkým v preparatívnej separácii rekombinantného proteínu. Svojimi vlastnosťami je vhodná aj pre purifikáciu rekombinantného proteínu z komplexnej proteínovej zmesi, ako v malej (angl. small-scale purification), tak aj vo veľkej mierke (angl. large-scale purification).
Rekombinantné proteíny sú cielene exprimované proteíny prostredníctvom nejakého expresného systému. Najčastejšie sú zložené z dvoch častí, preto im hovoríme fúzne proteíny:
- samotný čítací rámec proteínu
- proteínová kotva (tzv. proteínový prívesok, proteínový tag)
Proteínová kotva v podstate predstavuje len pokračovanie čítacieho rámca proteínu v podobe ďalších aminokyselín, ktoré vytvárajú určitý štruktúrny alebo funkčný motív, umožňujúci purifikáciu proteínu pomocou rôznych afinitných matríc. Proteínová kotva môže byť rôzne veľká (0,8-40 kDa). Vhodný purifikačný systém musí spĺňať niekoľko kritérií:
- proteínová kotva (spolu s proteínom) sa musí špecificky a reverzibilne viazať na vhodný nosič (matricu)
- purifikácia nesmie byť časovo ani technicky náročná
- výsledná kotva by nemala mať výrazný vplyv na celkový tvar (konformáciu) a funkciu proteínu, príp. by mala byť dodatočne odštiepiteľná vhodným enzýmom (proteázou)
| ligand matrice | proteínová kotva | veľkosť kotvy [kDa] | počet AK | poznámka |
| Peptidové kotvy | ||||
| napr. Ni2+, Zn2+, Co2+ | 6×His-tag | 0,8 | 6 | Metalo-chelátová afinitná chromatografia. Základná, technicky najmenej náročná a lacná purifikácia rekombinantného proteínu. Elučný roztok so >50 mM imidazolom. |
| protilátka | FLAG-tag | 1,01 | 8 | Imunoafinitná purifikácia. FLAG je jedna z prvých proteínových kotiev, ktorá bola patentovaná (1987). Elúcia pri pH = 3,0 alebo 2-5 mM EDTA. V prípade lokalizácie kotvy na C-terminálnom konci je dodatočne prirodzene oštiepiteľná pomocou enterokinázy, pretože predstavuje jej rozpoznávajúcu sekvenciu. |
| protilátka | T7-tag | 1,1 | 11 | Imunoafinitná purifikácia. T7-tag predstavuje sekvenciu z hlavného kapsidového proteínu bakteriofágu T7 a je súčasťou mnohých rekombinantných expresných vektorov využívajúcich T7 RNA-polymerázu. |
| protilátka | c-myc | 1,20 | 11 | Imunoafinitná purifikácia. Elúcia pomocou nízkeho pH. |
| protilátka | S-tag | 1,75 | 15 | Imunoafinitná purifikácia. S-tag je oligopeptid derivovaný z pankreatickej RNázy A. Môže zvyšovať solubilitu fúzneho proteínu, pretože obsahuje veľa polárnych a nabitých aminokyselín. |
| kalmodulín | kalmodulín-viažúci peptid (CBP) | 2,96 | 26 | Fúzne proteíny s CBP majú vyššiu solubilitu. CBP viaže kalmodulín na matrici pri neutrálnom pH a nízkej koncentrácii vápenatých katiónov. Elúcia taktiež prebieha pri neutrálnom pH, s prídavkom 2 mM EGTA, ktorá vychytáva Ca2+. |
| Proteínové kotvy | ||||
| glutatión | glutatión-S-transferáza (GST) | 26 | 211 | Podporuje solubilitu rekombinantného proteínu, efektívna, ale drahá purifikácia. Elúcia s 5-10 mM redukovaným glutatiónom (GSH). |
| amylóza | maltózu-viažúci proteín (MBP) | 40 | 396 | MBP je bakteriálny proteín, ktorý je zapojený do metabolizmu maltodextrínov (oligosacharidov). Proteínová kotva s MBP zvyšuje solubilitu fúzneho proteínu. Elúcia proteínov pomocou 10 mM maltózy. |
| - | tioredoxín E. coli (TrxA) | 11,7 | 109 | Potenciálne zvyšujú solubilitu fúzneho rekombinantného proteínu exprimovaného v bakteriálnom expresnom systéme. Neexistuje však pre ne vhodná, špecifická afinitná matrica. Z toho dôvodu je nutné použiť na ich selektívnu purifikáciu ďalšiu, menšiu peptidovú kotvu, napr. 6×His-tag. Keďže sú tieto proteínové kotvy veľké, je nutné ich odštiepiť pomocou špecifickej proteázy. |
| - | proteín-disulfid izomeráza I (DsbA) | 23,1 | 208 | Potenciálne zvyšujú solubilitu fúzneho rekombinantného proteínu exprimovaného v bakteriálnom expresnom systéme. Neexistuje však pre ne vhodná, špecifická afinitná matrica. Z toho dôvodu je nutné použiť na ich selektívnu purifikáciu ďalšiu, menšiu peptidovú kotvu, napr. 6×His-tag. Keďže sú tieto proteínové kotvy veľké, je nutné ich odštiepiť pomocou špecifickej proteázy. |
| - | transkripčný terminátor-antiterminátor fágu λ (NusA) | 54,9 | 495 | Potenciálne zvyšujú solubilitu fúzneho rekombinantného proteínu exprimovaného v bakteriálnom expresnom systéme. Neexistuje však pre ne vhodná, špecifická afinitná matrica. Z toho dôvodu je nutné použiť na ich selektívnu purifikáciu ďalšiu, menšiu peptidovú kotvu, napr. 6×His-tag. Keďže sú tieto proteínové kotvy veľké, je nutné ich odštiepiť pomocou špecifickej proteázy. |
Zdroj:
- Pančík P.: M3 proteín Myšieho herpetického vírusu, nový modulátor imunitnej odpovede. (2013). Virologický ústav Slovenskej akadémie vied, Prírodovedecká fakulta UK, Bratislava. Dizertačná práca.
- Page R.: Strategies for Maximizing Heterologous Protein Expression in E. coli with Minimal Cost. (2007). Department of Molecular Biology, Biochemistry and Cell Biology, Brown University, Providence, Rhode Island
- Protein tag - Wikipedia, Free Encyclopedia
Biotín predstavuje osobitnú skupinu značenia proteínov. Biotín je známy ako vitamín H, ktorý zohráva úlohu pri metabolizme mastných kyselín, aminokyselín a glukózy. Má široké uplatnenie v biotechnológiách, pretože zabezpečuje veľmi pevnú, špecifickú väzbu s ligandom, ktorým je streptavidín (avidín). Značenie proteínov biotínom sa nazýva biotinylácia. Purifikácia biotinylovaných proteínov si však často vyžaduje elúciu za denaturujúcich podmienok (6M GuHCl, pH = 1,5), pretože je táto väzba veľmi pevná. Na druhej strane, malá veľkosť biotínu (244 Da) málokedy vyžaduje jeho dodatočné odštiepenie z rekombinantného proteínu.
V niektorých prípadoch je potrebné proteínovú kotvu odštiepiť, aby bolo možné obnoviť biologickú aktivitu exprimovaného rekombinantného proteínu. Toto štiepenie zabezpečujú špeciálne proteázy, ktoré rozoznávajú určitý sekvenčný alebo štruktúrny motív. Tieto sekvencie sú spravidla súčasťou expresných vektorov (napr. bakteriálny expresný systém pET).
| meno | pôvod | typ | rozpoznávajúca sekvencia [1-písmenový kód] |
| HRV 3C | ľudský vírus nádchy | cysteínová proteáza | LEVLFQG▼P |
| enterokináza | ľudský tráviaci enzým z dvanástnika | serínová proteáza | DDDDK▼X |
| TEV | Tobacco Etch Virus | cysteínová proteáza | ENLYFQG▼S |
| faktor X | koagulačný faktor X | serínová proteáza | I(E/D)GR▼ |
| trombín | koagulačný faktor II | serínová proteáza | LVPA▼GS |
| TAGZyme | DAPáza (+ Qcykláza) | exopeptidáza | špecifická pre 6×His-tag (firma Qiagen) |
| SUMO | ubikvitínu-podobná proteáza | cysteínová proteáza | rozoznáva štruktúrny motív |
Zdroj:
- Protein Expression Facility, University of Queensland, Australia
- Lebendiker M.: Cleavage Sites Table for Fusion Proteins. (2002). Protein Purification Facility. Wolfson Centre for Applied Structural Biology, The Hebrew University of Jerusalem, Izrael
Z tabuľky vyplýva, že napriek enzymatickému odštiepeniu proteínovej kotvy ostáva niekoľko aminokyselín, tvoriach rozpoznávajúce miesto pre proteázu, stále pripojených k rekombinantnému proteínu, čo môže, ale nemusí ovplyvňovať jeho biologickú aktivitu. Zároveň treba dbať na prípadné prečistenie proteínovej zmesi od nepoštiepených molekúl, pretože tento proces nie je nikdy efektívny na 100% (napr. v prípade enterokinázy závisí od prvej aminokyseliny X v pozícii za štiepnym miestom). Dizajn rekombinantného proteínu a postupu jeho purifikácie by sa tak mal predovšetkým opierať o menšie kotvy (napr. 6×His-tag, biotín).
Metalo-chelátová afinitná chromatografia link
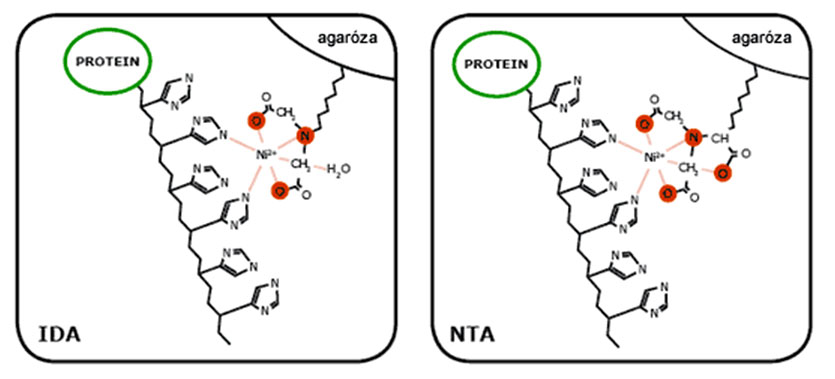
Technika metalo-chelátovej afinitnej chromatografie (angl. immobilized metal affinity chromatography, IMAC) vychádza z interakcie histidínovej peptidovej kotvy (6×His-tag) s matricou, ktorú tvoria 3 zložky:
- polysacharidový nosič - najčastejšie agaróza (napr. Sepharose CL-6B pri Ni-NTA Agarose od firmy Qiagen)
- kyselina imino-dioctová (IDA) alebo kyselina nitrilo-trioctová (NTA)
- chelatačný ligand, ktorý interaguje s postrannými reťazcami aminokyseliny histidínu - dvojmocný katión kovového prvku (najčastejšie Ni2+)
Histidínová kotva sa môže nachádzať na C- alebo N-terminálnom konci rekombinantného proteínu, alebo kdekoľvek v jeho čítacom rámci, nemala by však ovplyvniť jeho natívnu konformáciu a biologickú aktivitu. Napriek veľkej špecificite matrice k tejto peptidovej kotve, často dochádza ku kopurifikácii kontaminujúcich proteínov. Jej rozsah súvisí predovšetkým s chelatačným potenciálom kovového prvku ale aj samotnou matricou. Vzhľadom na ión kovu, klesá špecificita (a vzrastá množstvo kopurifikujúcich, kontaminujúcich proteínov) približne takto:
Ni2+ > Co2+ > Zn2+ > Cu2+
NTA je špecifickejšia v tom zmysle, že tvorí o jednu koordinačnú väzbu s kovovým prvkom naviac, v porovnaní s IDA. To môže mať za následok nižší výťažok, ale o to špecifickejšiu interakciu s rekombinantným proteínom.
Elúcia prebieha najčastejšie použitím kompetitívneho ligandu, ktorým je imidazol v koncentrácii väčšinou >50 mmol/l. Pri purifikácii proteínov sa nesmie použiť EDTA, ani iné podobné chelatačné činidlo, pretože by vyviazalo kovový prvok z matrice a separácia by bola neúspešná.