Autor: Peter Pančík
Publikované dňa:
Citácia: PANČÍK, Peter. 2016. Biopedia.sk: Regulácia génovej expresie. [cit. 2024-04-24]. Dostupné na internete: <https://biopedia.sk/molekularna-biologia/regulacia-genovej-expresie>.
Génová expresia (sloveso exprimovať) predstavuje vyjadrenie genetickej informácie úseku nukleovej kyseliny (DNA, RNA u RNA-vírusov) - génu - do podoby finálneho produktu, ktorým je bielkovina alebo funkčná RNA (napr. tRNA, rRNA). Niekedy sa zvykne nahrádzať pojmom génová transkripcia, tá však vo väčšine prípadov predstavuje len časť génovej expresie, pretože mnohé primárne transkripty ešte podliehajú post-transkripčným úpravám a u štruktúrnych génov aj translácii a post-translačným úpravám. Génová expresia teda zahŕňa všetky procesy, ktoré začínajú transkripciou génu a končia tvorbou hotového produktu schopného vykonávať svoju biologickú funkciu. Ak o expresii génu hovoríme z pohľadu štúdia proteomiky, čo je veda o štruktúre a funkcii proteínov a využívajúca produkciu heterologických rekombinantných proteínov, zvykne sa používať aj pojem expresia proteínu.
Najčastejšie je termín génová expresia spojený s vyjadrením aktivity génu, čo možno voľne preložiť ako frekvenciu, resp. mieru, s akou (a či vôbec) dochádza k transkripcii génu a vzniku funkčného proteínu. Čím je aktivita, resp. expresia génu vyššia, tým viac proteínu bude v bunke možné detegovať, a to pomocou rôznych molekulárnych metód (napr. SDS-PAGE). Môžeme teda relatívne porovnávať hladiny génovej expresie rôznych génov u dvoch rôznych bunkových línií alebo sledovať efekt vírusovej infekcie, príp. terapeutického účinku látok, na expresiu vybraných génov pred a po ovplyvnení buniek (vírusom, chemickou látkou), a pod.
Každá bunka vlastní mechanizmy, ktoré ovplyvňujú génovú expresiu na každej úrovni:
- transkripcia,
- posttranskripčné úpravy,
- transport cez jadrovú membránu (u eukaryotov),
- translácia,
- posttranslačné úpravy.
Najdôležitejšia je pritom regulácia na úrovni génovej transkripcie. Regulácia génovej expresie súvisí nielen s ovplyvnením množstva finálneho produktu, ale najmä so správnym načasovaním jeho prítomnosti v bunke počas jej života. Je teda základným predpokladom bunkovej diferenciácie, morfogenézy a evolučnej adaptability každého živého organizmu.
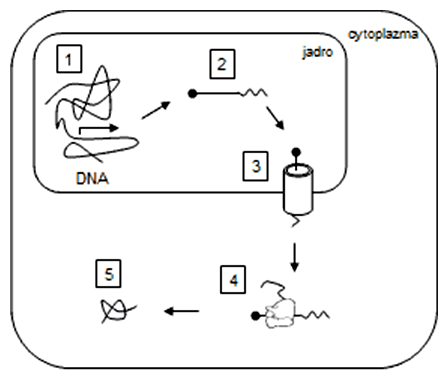
Regulácia génovej expresie u prokaryot link
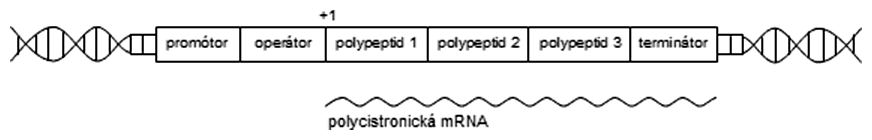
Ako je známe, mnohé molekuly v bunke vznikajú chemickou premenou iných molekúl. Deje sa to za účasti viacerých enzýmov, pričom produkt jedného enzýmu je substrátom pre druhý enzým atď. Takáto kaskáda enzymatických reakcií, prostredníctvom ktorých vznikne z východzej chemickej molekuly výsledný produkt, sa nazýva enzymatická dráha. Všetky enzýmy sú bielkovinovej povahy, a teda kódované génmi. U baktérií a mnohých vírusov často dochádzalo k zoskupovaniu génov, ktorých produkty - enzýmy - patria do enzymatickej dráhy, aby bola na určitý podnet (napr. prítomnosť substrátu v živnom médiu) zahájená transkripcia všetkých génov naraz, pričom tieto oblasti dostali názov polycistronické transkripčné jednotky. Polycistronické mRNA sú typické pre všetky prokaryoty (baktérie, archeóny), ale naopak takmer úplne chýbajú u eukaryot. Každý gén, ktorý je súčasťou polycistronickej transkripčnej jednotky, má vlastnú ribozomálnu väzobnú sekvenciu (RBS, angl. ribosome binding site) nazývanú Shine-Dalgarnova sekvencia.
U baktérií predstavuje ovplyvnenie génovej transkripcie kľúčový spôsob regulácie génovej expresie. Signál k zahájeniu alebo zastaveniu transkripcie prichádza väčšinou v podobe malej chemickej molekuly. Táto molekula však často nemá chemické predpoklady, aby mohla interagovať s DNA a nejakým spôsobom spustiť transkripciu. Využíva k tomu proteín, ktorý má minimálne dve špecifické funkčné oblasti, tzv. väzbové domény: jednou viaže spomenutú molekulu a druhou DNA. Proteín s takýmito vlastnosťami, ktorý dokáže regulovať transkripciu, sa nazýva regulačný proteín. Na základe charakteru spomenutej chemickej molekuly a regulačného proteínu rozlišujeme pozitívnu a negatívnu reguláciu. Transkripčná jednotka však môže byť zároveň regulovaná oboma spôsobmi.
Negatívna regulácia operónu link
Regulovateľná transkripčná jednotka, ktorá je prepisovaná na určitý signál, sa nazýva u baktérii operón. Okrem promótora, nevyhnutného pre väzbu RNA-polymerázy, a jedného alebo viacerých génov obsahuje aj regulačnú oblasť nazývanú operátor. Operátor je sekvencia DNA, na ktorú sa viaže regulačný proteín, nazývajúci sa v prípade negatívnej regulácie represor. Represor je teda negatívny regulačný proteín v tom zmysle, že jeho väzba na DNA v oblasti operátora znemožňuje RNA-polymeráze transkripciu. Represor je aktívny, ak je naviazaný na operátore.
Enzýmová indukcia link
Enzýmová indukcia je regulovateľná syntéza enzýmov enzymatickej dráhy, ktorá je zahájená v prítomnosti chemickej zlúčeniny - induktora. Induktor sa viaže na pôvodne aktívny represor, čím spôsobí jeho vyviazanie z operátora (odpadnutie od DNA), a tým umožní RNA-polymeráze prepísať danú transkripčnú jednotku. Transkripčná jednotka, ktorá je regulovaná takýmto spôsobom, sa nazýva indukovateľný operón. Príkladom je syntéza enzýmov laktózového operónu, ktoré metabolizujú laktózu na glukózu. Tieto enzýmy sa však začnú tvoriť len v prípade, ak sa laktóza vyskytne v živnom médiu. Laktóza spôsobuje vyviazanie represora z operátora, takže je zároveň induktorom laktózového operónu. Ak sa laktóza z média vyčerpá, začne chýbať aj ako induktor laktózového operónu a syntéza enzýmov sa zastaví.
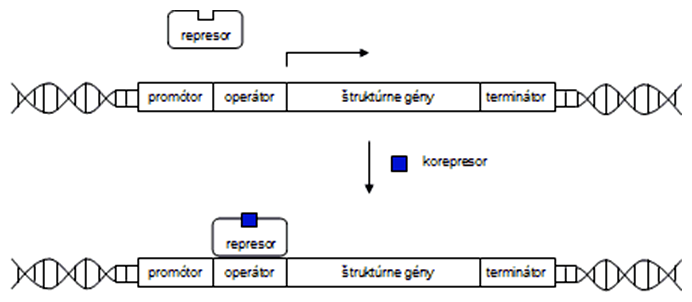
Enzýmová represia link
Enzýmová represia je opakom enzýmovej indukcie. Vyskytuje sa u operónov, ktoré obsahujú gény kódujúce enzýmy enzymatickej dráhy pre syntézu aminokyselín. Princíp je taký, že sú prepisované dovtedy, kým kritické nahromadenie konečného produktu enzymatickej dráhy (aminokyseliny) nespôsobí zastavenie ich transkripcie. Aminokyselina v tomto prípade funguje ako korepresor, ktorý väzbou s represorom (dovtedy inaktívnym) spôsobí jeho naviazanie na operátor a zastavenie transkripcie. Transkripčná jednotka, ktorá je regulovaná prítomnosťou korepresora, sa nazýva reprimovateľný operón. Príkladom je tryptofánový alebo histidínový operón.

Pozitívna regulácia operónu link
Pozitívna regulácia operónu zahŕňa väzbu regulačného proteínu - aktivátora - na DNA. Zvyčajne sa viaže na iné miesto ako operátor (napr. na oblasť promótoru alebo upstream od promótora). Aktivátor je pozitívny regulačný proteín v tom zmysle, že jeho väzba na DNA umožňuje transkripciu. Pozitívna regulácia je podstatou katabolickej represie. Reguláciou tohto typu sú charakteristické operóny, ktoré kódujú proteíny enzymatických dráh spracúvajúcich (katabolizujúcich) cukry.
Laktózový operón link
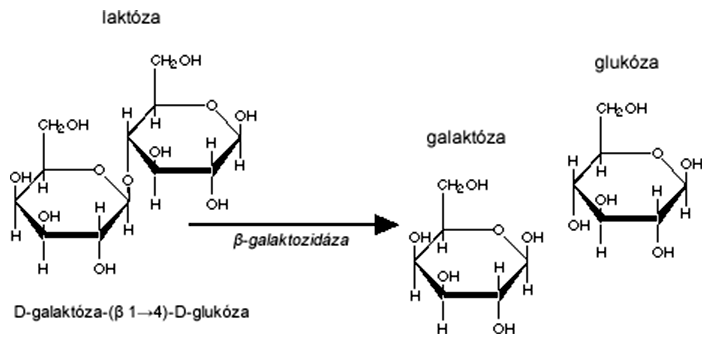
Laktóza je disacharid zložený z galaktózy a glukózy, ktoré sú spojené β(1-4)-glykozidovou väzbou. Baktérie (napr. Escherichia coli), ktoré majú laktózový operón (skrátene lac-operón), dokážu tento cukor metabolizovať. Súčasťou lac-operónu sú 3 gény, z ktorých najdôležitejší je lacZ kódujúci enzým β-galaktozidázu (hydrolyzuje glykozidovú väzbu). Všetky tri gény sú súčasťou jednej polycistronickej mRNA, a teda sú prepisované súčasne.
Lac-operón je príkladom operónu, ktorý je regulovaný negatívne aj pozitívne. Negatívnu reguláciu zahŕňa represor, ktorý je produktom génu lacI. Tento gén sa nachádza mimo laktózového operónu. LacI-represor (proteín) v aktívnom stave zabraňuje transkripcii lac-operónu v neprítomnosti laktózy. Ak je laktóza pridaná do živného média, pôsobí ako induktor, ktorý sa viaže na lacI-represor, čím sa tento vyviaže z operátora. Tým je splnená jedna z dvoch podmienok expresie laktózového operónu. Druhou podmienkou je prítomnosť aktivátora, ktorý sa viaže na DNA upstream od promótora. Pozitívny regulačný proteín s označením CAP (angl. katabolický aktivačný proteín) vytvorí komplex s cAMP (cyklický adenozínmonofosfát) a umožní tak RNA-polymeráze iniciovať transkripciu. Dôležité pritom je, že ak je v prostredí prítomná s laktózou zároveň aj glukóza, je pre bunku výhodnejšie využiť glukózu namiesto laktózy, kvôli ktorej treba mrhať energiu a živiny na syntézu enzýmov. Prítomnosť glukózy spôsobuje, že klesá koncentrácia cAMP, čím nevzniká aktívny komplex CAP.cAMP a transkripcia lac-operónu nebeží. Glukóza je zároveň produktom katabolizmu laktózy, preto sa táto regulácia nazýva katabolická represia alebo glukózový efekt.

Tryptofánový operón link
Tryptofánový operón (skrátene trp-operón) kóduje enzýmy zapojené do syntézy aminokyseliny tryptofánu. Trp-operón je regulovaný negatívne prostredníctvom represora kódovaného génom trpR nachádzajúcim sa, podobne ako lacI, na inom mieste bakteriálneho chromozómu. Korepresorom je tryptofán, ktorý sa pri nadbytku viaže na represor a zastavuje expresiu tryptofánového operónu.
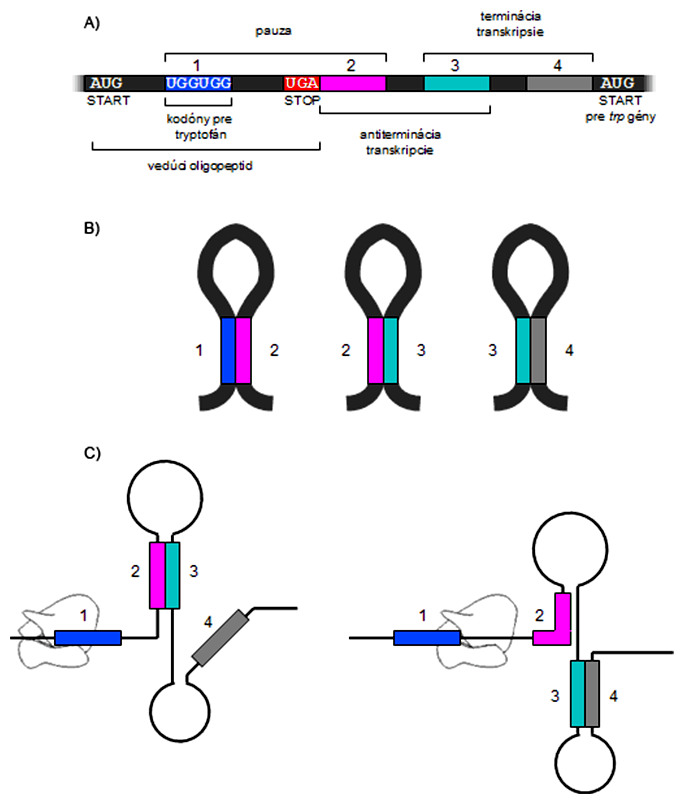
Okrem tejto regulácie je však trp-operón regulovaný aj prostredníctvom tzv. atenuácie, pretože represor je v tomto prípade nedostačujúci, a RNA-polymeráza mnohokrát "prekĺzne" aj v prítomnosti tryptofánu. Princíp atenuácie spočíva v úzkom prepojení bakteriálnej transkripcie s transláciou a v tvorbe sekundárnych štruktúr - vláseniek - na mRNA. Začiatok mRNA trp-operónu kóduje krátky oligopeptid, pričom pri jej konci sa nachádza dvojica kodónov pre tryptofán. To je veľmi nezvyčajné, pretože tryptofán je vo všeobecnosti vzácna aminokyselina. Ich prítomnosť je nevyhnutná pre správne fungovanie atenuácie.
Veľmi skoro po začiatku transkripcie trp-operónu sa na vedúcu sekvenciu mRNA pripája ribozóm, pričom RNA-polymeráza ešte len prepisuje prvý gén trp-operónu. Párovanie vláseniek 1-2 spôsobí krátke zbrzdenie RNA-polymerázy - pauzu - pri iniciácii translácie dovtedy, kým ribozóm nenasadne na mRNA, potom sa vlásenka 1-2 rozpadá. Ďalší priebeh závisí od koncentrácie tryptofánu v bunke.
Ak je tryptofánu dostatok, ribozóm preloží celý oligopeptid a mRNA tým pádom vytvorí vlásenky medzi regiónmi 3-4, čo destabilizuje RNA-polymerázu a ukončí transkripciu termináciou. Ak je nedostatok tryptofánu, ribozóm zastane na kodónoch pre tryptofán (pretože je v bunke veľmi málo aktivovaných tRNATrp), takže sa, na druhej strane, stihne vytvoriť vlásenka medzi oblasťami 2-3, čo zabráni vytvoreniu terminátorovej vlásenky (t.j. 3-4) - antiterminácia. Pri atenuácii teda rozhoduje rýchlosť, akou ribozóm postupuje po mediátorovej RNA, a tá závisí od koncentrácie kritickej aminokyseliny. Podobný mechanizmus reguluje aj syntézu histidínu, fenylalanínu, valínu, leucínu a izoleucínu.
Alternatívne σ-faktory link
σ-faktor (sigma-faktor) je kofaktor (proteín) viažúci sa jednak so špecifickou sekvenciou promótora a jednak s RNA-polymerázou. Za určitých podmienok môže u baktérií dochádzať k tvorbe σ-faktorov, ktoré rozoznávajú inú konvenčnú sekvenciu ako TTGACAT, čo vedie k zvýšeniu väzby (afinity) RNA-polymerázy k dovtedy neprepisovaným operónom, resp. transkripčným jednotkám s odlišnou sekvenciou promótora.
Regulácia génovej expresie u eukaryot link
Baktériám slúži regulácia génovej expresie predovšetkým na prispôsobenie sa podmienkam prostredia, takže bunky dokážu optimalizovať svoj rast a delenie. U mnohobunkových eukaryotických organizmov je prvoradý účel kontroly génovej expresie regulácia embryonálneho vývinu a realizácia špecializovaných genetických programov, ktorými sú charakteristické rôzne diferencované bunky. Pritom ak už bunka nastúpi na cestu diferenciácie v určitý bunkový typ, tieto zmeny sú nevratné, čo je rozdiel oproti flexibilnej bakteriálnej enzýmovej indukcii a represii. U eukaryot rozhoduje často o diferenciácii buniek len jeden kľúčový signál, ktorý spúšťa kaskádu mnohých reakcií vedúcich k bunkovej špecializácii.
Eukaryotický chromatín má okrem organizovania DNA do kompaktných, ľahko segregovateľných štruktúr - chromozómov - počas mitotického delenia buniek aj významnú regulačnú funkciu. Oblasti s kondenzovaným chromatínom (heterochromatínom) nie sú prístupné regulačným proteínom a RNA-polymerázam, takže gény v týchto miestach nie sú exprimované. To znamená, že eukaryoty nepotrebujú prokaryotické operóny, ktoré by upravovali (modulovali) transkripciu. Namiesto toho existujú špeciálne proteíny, ktoré podľa potreby zabezpečujú remodeláciu chromatínu, čím sprístupňujú promótory regulačným proteínom. Z uvedeného vyplýva, že euchromatín je transkripčne aktívny, zatiaľčo heterochromatín je transkripčne inaktívny. Prokaryotické operóny často pozostávajú z viacerých génov, ktoré sa transkribujú spoločne. U eukaryot sú polycistronické transkripčné jednotky skôr výnimkou, napriek tomu je však expresia funkčne dopĺňajúcich sa enzýmov enzymatických dráh koordinovaná.
Dôležitú úlohu v regulácii génovej expresie u eukaryot má štruktúra promótora, ktorý okrem základných sekvencií funkčne zrovnateľných s prokaryotickými obsahuje veľmi často tzv. enhancery, čo možno preložiť ako zosilovače transkripcie. Na ne sa viažu špecifické regulačné proteíny, ktoré zvyšujú expresiu príslušných génov. Tieto sekvencie sa môžu nachádzať aj niekoľko tisíc kilobáz mimo génu, ktorého transkripciu modulujú. Ich pôsobenie je umožnené ohybom DNA pomocou regulačných proteínov, ktoré ich priestorovo prepoja s miestom začiatku transkripcie. Podobné elementy - silencery naopak zoslabujú transkripciu. V mnohých prípadoch môže byť transkripcia z génu zoslabená aj metyláciou DNA (pridávanie metylovej CH3- skupiny, najčastejšie na cytozín) v promótorovej oblasti. Metylované cytozínové zvyšky nie sú rozoznávané regulačnými proteínmi. Kombináciou rôznych enhancerov, silencerov a proteínov viažúcich základné promótorové sekvencie je umožnená dôsledná kontrola množstva produktu na úrovni transkripcie.
V časovej regulácii génovej expresie u eukaryot majú veľký význam hormóny. Tie spúšťajú signálne dráhy vedúce často k aktivácii špecifických regulačných proteínov, ktoré viažu DNA a ovplyvňujú transkripciu. Najpriamejšie ovplyvňujú transkripciu steroidné hormóny (testosterón, progesterón, aldosterón), ktoré vstupujú do buniek a priamo sa viažu na konkrétne aktivátory príslušných génov. Veľmi dôležitá je pri mnohobunkových organizmoch bunková komunikácia, ktorá prebieha pomocou rôznych receptorov a kontaktných molekúl. Asi najkomplexnejšie je regulovaný priebeh bunkového cyklu, ktorý pozostáva z mnohých fáz a viacerých kontrolných uzlov. Každá fáza a každá bunková odpoveď na vonkajšie prostredie je vždy spojená s aktiváciou alebo naopak represiou špecifických génov.
Z regulácie génovej expresie eukaryot na ďalších úrovniach spomeniem interferujúce RNA (RNAi), ktoré využívajú aj mnohé prokaryoty. Je to pomerne novo objavená skupina funkčných RNA, ktoré negatívne regulujú génovú expresiu na úrovni RNA produktu, resp. mRNA. Sú to malé molekuly RNA, ktoré znižujú množstvo vyprodukovanej mRNA tým, že sú k nej komplementárne, čím sa prechodne vytvorí dvojvláknová RNA. Tá je potom signálom pre degradáciu molekuly mRNA. Na úrovni RNA existuje ešte regulácia v podobe rôznych stabilizujúcich alebo naopak destabilizujúcich proteínov, ktoré ovplyvňujú post-transkripčné úpravy, príp. transport RNA do cytoplazmy cez jadrové póry.