Autor: Peter Pančík
Publikované dňa:
Citácia: PANČÍK, Peter. 2016. Biopedia.sk: Transkripcia. [cit. 2024-04-17]. Dostupné na internete: <https://biopedia.sk/molekularna-biologia/transkripcia>.
Transkripcia predstavuje prepis genetickej informácie z jedného typu nukleovej kyseliny do druhého typu. Štandardne sa tým myslí prepis informácie z DNA do RNA. Reverzná transkripcia, ktorou je charakteristický životný cyklus niektorých RNA vírusov, a špeciálne polymerázy - telomerázy, predstavuje opačný smer, t.j. podľa predlohy v podobe RNA sa syntetizuje DNA. Zostaňme však pri štandardnom ponímaní termínu transkripcie. Podobný pojem je génová expresia (doslova "vyjadrenie génu", resp. jeho genetickej informácie), ktorý predstavuje kvantitatívnu mieru transkripcie. Gény s vysokou génovou expresiou podliehajú transkripcii často a vo väčšej intenzite ako gény s nízkou génovou expresiou. Hlavný enzým podieľajúci sa na transkripcii je RNA-polymeráza.
Rozlišujeme niekoľko typov produktov transkripcie:
| typ RNA | anglický názov | skratka | miesto syntézy |
| mediátorová RNA | messenger RNA | mRNA | jadro |
| transferová RNA | transfer RNA | tRNA | jadro |
| ribozomálna RNA | ribosomal RNA | rRNA | hlavne jadierko |
| malé jadrové RNA | small nuclear RNA | snRNA | jadro |
| malé jadierkové RNA | small nucleolar RNA | snoRNA | jadierko |
Okrem mRNA sa ostatné typy RNA molekúl zvyknú nazývať termínom nekódujúce alebo funkčné RNA, pretože nie sú rozoznávané ribozómom a nie sú podľa genetického kódu prekladané do sekvencie proteínu, čiže vykonávajú svoju funkciu ako molekuly RNA. RNA reťazec, ktorý je syntetizovaný RNA-polymerázou, sa označuje pojmom transkript.
Všeobecná štruktúra génu link
Gén predstavuje kompletnú sekvenciu nukleovej kyseliny (DNA alebo RNA), ktorá nesie informáciu pre syntézu určitého produktu (funkčnej RNA alebo proteínu). Gény, ktoré sú prepisované do mRNA a následne sa podľa tejto mRNA syntetizujú proteíny, označujeme pojmom štruktúrne gény.
Nie celý gén podlieha transkripcii. Prvý nukleotid na DNA, ktorý sa prepíše do sekvencie RNA, sa označuje +1. Pozícia na sekvencii DNA, ktorá sa nachádza pred týmto nukleotidom, má záporné číslovanie, pričom 0 neexistuje (napr. -20 je nukleotid umiestnený dvadsať miest pred pozíciou +1). Sekvencie nachádzajúce sa pred +1 sú označované ako upstream (angl. hore prúdom), za týmto nukleotidom sú downstream (angl. dole prúdom), pričom toto pozicovanie je relatívne vzhľadom na nami vybranú pozíciu (v našom príklade nukleotid +1).
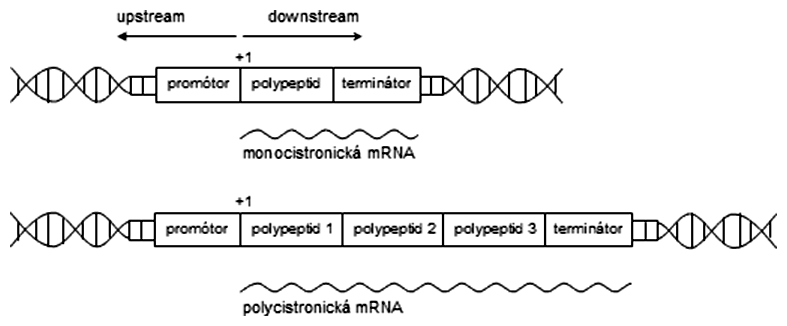
Trochu odlišný pojem od génu je transkripčná jednotka. Je to úsek DNA, z ktorého sa syntetizuje jedna súvislá molekula RNA. Jedna transkripčná jednotka môže tak pozostávať z viacerých génov. To je veľmi častý prípad u prokaryot, u ktorých súvisí s ekonomikou syntézy, pretože gény pre syntézu viacerých enzýmov postupne metabolizujúcich určitú živinu môžu byť usporiadané za sebou do jednej transkripčnej jednotky a tak prepisované na určitý signál (prítomnosť živiny v prostredí) spoločne. Takýto transkript, ktorý obsahuje informáciu pre syntézu viacerých polypeptidových reťazcov (proteínov), sa označuje pojmom polycistronická mRNA. U eukaryot sa stretávame väčšinou s monocistronickou mRNA, to znamená, že jedna molekula RNA obsahuje informáciu pre syntézu len jedného polypeptidového reťazca (proteínu).
Najdôležitejšou súčasťou každého génu, resp. transkripčnej jednotky je promótor. Je to úsek na DNA, ktorý priamo súvisí s rozoznávaním génu RNA-polymerázou a iniciáciou transkripcie. Každý promótor má viaceré funkčné oblasti, ktorých umiestnenie a nukleotidová sekvencia je odlišná u prokaryot a eukaryot, ale aj v rámci jednej či druhej skupiny nie sú tieto sekvencie úplne rovnaké. Nukleotidová sekvencia, ktorá sa spomedzi ostatných sekvencií majúcich rovnakú funkciu vyskytuje najčastejšie, sa označuje ako konvenčná sekvencia (angl. consensus sequence). Konkrétne funkčné oblasti ako aj ich konvenčné sekvencie spomeniem osobitne pri charakteristike prokaryotickej a eukaryotickej transkripcie nižšie v tomto texte.
Všeobecný priebeh transkripcie link
Priebeh transkripcie môžeme podobne ako replikáciu DNA rozdeliť na 3 fázy, ktoré sú principiálne u prokaryot a eukaryot takéto:
Iniciácia transkripcie link
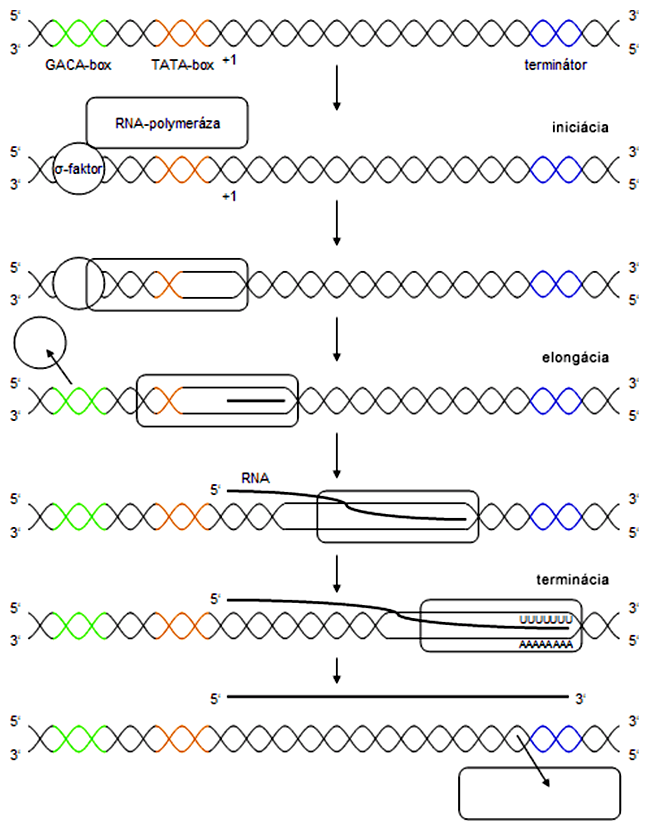
Zahŕňa naviazanie RNA-polymerázy na promótor, ktorý predstavuje presne sekvenčne definovanú oblasť na DNA. Prokaryotický a eukaryotický promótor sa od seba odlišujú zložitosťou aj primárnou štruktúrou. Iniciačná fáza transkripcie končí zahájením syntézy reťazca RNA v smere 5'-3' podľa komplementarity báz len na jednom vlákne opačnej polarity - 3',5', pre ktoré sa používajú pomenovania matricový reťazec (angl. template strand) (Russell, Genetics, 2006), templátový reťazec (angl. template strand) (Lodish, Molecular Cell Biology, 2003) alebo negatívny reťazec (Rosypal, Úvod do molekulární biologie II., 1999). RNA-polymeráza nevyžaduje pre svoju funkciu primer ako substrát.
Elongácia transkripcie link
RNA-polymeráza postupne pridáva nukleozid-5'-monofosfáty na 3'-koniec reťazca RNA, pričom sa posúva po kódujúcom reťazci DNA, ktorého smer fosfodiesterových väzieb je 3'-5'. Celú molekulu RNA transkriptu syntetizuje jedna molekula RNA-polymerázy. Na rozdiel od replikácie DNA, gén môže byť (a vždy aj je) prepisovaný viackrát, zatiaľčo DNA-polymeráza zreplikuje každý úsek DNA len raz. Počas transkripcie sú potrebné aj topoizomerázy, pretože RNA-polymeráza pred sebou vytvára pozitívne superhelixy (previnutú DNA) a za sebou negatívne (podvinutú DNA).
Terminácia transkripcie link
Zastavenie elongácie reťazca RNA na konci génu a vyviazanie RNA-polymerázy z matricového reťazca DNA.
Mnohé transkripty, obzvlášť eukaryotické, sú ďalej upravované (modifikované). Tieto procesy označujeme ako post-transkripčné úpravy. Vtedy sa pôvodná RNA zvykne nazývať primárnym transkriptom a post-transkripčne upravená RNA ako maturovaná RNA.
Úloha RNA-polymeráz link
RNA-polymerázy sú enzýmy, ktoré katalyzujú syntézu jednovláknového reťazca RNA z voľných nukleozid-5'-trifosfátov, ktoré pripájajú na 3'-OH koniec syntetizujúceho sa RNA reťazca ako monofosfáty, pričom ako templát využívajú kódujúce vlákno DNA s polaritou 3',5'. Podobne ako DNA-polymerázy, dokážu polymerizovať reťazec len v smere 5'-3'. Na rozdiel od replikácie však nevyžadujú primer na začatie transkripcie. RNA-polymeráza má v porovnaní s DNA-polymerázou veľmi nízku 3',5'-exonukleázovú aktivitu (proof-reading aktivitu), čo znamená, že počas elongácie RNA nie vždy vyštepuje nesprávne zaradený nukleotid. To však nie je veľká tragédia, pretože RNA neslúži na uchovanie genetickej informácie (zatiaľčo DNA pri replikácii áno), a okrem toho gén je prepisovaný v mnohých kópiách, takže občasná chyba v niektorých transkriptoch RNA neovplyvní celkovú funkciu.
Prokaryoty majú len jeden druh RNA-polymerázy, ktorá katalyzuje syntézu RNA zo všetkých génov.
Eukaryoty naopak používajú viacero typov RNA-polymeráz, ktoré zabezpečujú transkripciu z rôznych génových skupín:
- RNA-polymeráza I - vyskytuje sa v jadierku, transkribuje gény pre rRNA
- RNA-polymeráza II - vyskytuje sa v jadre, transkribuje štruktúrne gény a gény pre niektoré malé RNA
- RNA-polymeráza III - vyskytuje sa v jadre, transkribuje gény pre tRNA, 5S-rRNA a niektoré malé RNA
Prokaryotická transkripcia link
Prokaryotická transkripcia je v mnohom jednoduchšia ako eukaryotická. Prokaryotický promótor má nasledovné funkčné oblasti:
- GACA-box - konvenčná sekvencia 5'-TTGACAT-3', pozícia ~(-35)
- TATA-box (Pribnowov box) - konvenčná sekvencia 5'-TATAAT-3', pozícia ~(-10)
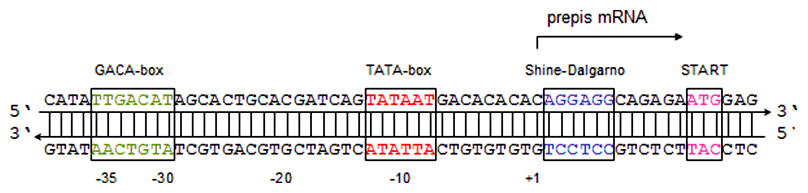
U prokaryotov existuje len jeden typ RNA-polymerázy, ktorá priamo rozoznáva GACA-box a bez pomoci ostatných enzýmov katalyzuje odvíjanie dvojzávitnice DNA, ktoré začína v mieste TATA-boxu. Promótor je rozoznávaný pomocou proteínovej podjednotky RNA-polymerázy, tzv. σ-faktoru (sigma-faktoru), ktorý sa po iniciácii transkripcie uvoľní, čo "odštartuje" elongáciu. Syntetizovaný reťazec RNA netvorí trvalé dvojvlákno s matricovým reťazcom DNA, na rozdiel od dcérskeho a rodičovského reťazca DNA pri replikácii, ale je z matricového reťazca vytesnený krátko po prechode RNA-polymerázy. V priebehu elongácie sa dvojzávitnica pred polymerázou odvíja a po pohybe enzýmu opäť spája, pričom k tomu nie sú potrebné žiadne prídavné enzýmy.
Terminácia transkripcie končí v mieste terminátora. Rozoznávame v podstate dva spôsoby, ako môže dôjsť k ukončeniu transkripcie. Prvý súvisí so syntézou špecifickej, na uracil bohatej sekvencie RNA (terminátorová sekvencia na DNA je v tomto mieste tvorená niekoľkými za sebou idúcimi adenínmi), ktorá labilizuje spojenie RNA-DNA v mieste pohybu polymerázy (pretože spojenie A-U je slabšie ako C-G). To má za následok samovoľné vyviazanie enzýmu z DNA a ukončenie transkripcie. Druhý spôsob terminácie využíva pomocný proteín - ρ-faktor (ró-faktor), ktorý katalyzuje vyviazanie RNA-polymerázy v mieste terminátorovej sekvencie, ktorá je odlišná od predošlého zloženia (neobsahuje opakovania A).
Keďže prokaryotická transkripcia prebieha v podstate v cytoplazme, okamžite je v prípade transkripcie štruktúrnych génov nasledovaná transláciou. Vyplýva z toho, že žiadne post-transkripčné úpravy, ktoré súvisia s transportom transkriptu cez jadrovú membránu, nie sú u prokaryot potrebné. Stratégia prokaryotickej proteosyntézy ako aj zloženie transkripčných jednotiek sú založené na víťazstve najrýchlejšie sa množiaceho, bez zbytočnej spotreby látok a energie.
Eukaryotická transkripcia link
Keďže hlavný dôraz je kladený na proteosyntézu, zameriam sa na popis promótora štruktúrnych génov a priebeh transkripcie katalyzovanej RNA-polymerázou II. Eukaryotický promótor má viacero funkčných oblastí, z ktorých najdôležitejšie sú:
- GC-box - konvenčná sekvencia 5'-GGGCGG-3', pozícia ~(-90)
- CAAT-box (čítaj 'kæt box' - ako mačka po anglicky) - konvenčná sekvencia 5'-GGCCAATCT-3', pozícia ~(-80)
- TATA-box (Hognessov box) - konvenčná sekvencia 5'-TATAAAA-3', pozícia ~(-30)
Eukaryotickej transkripcie sa zúčastňuje viacero pomocných proteínov, ktoré sa súhrnne nazývajú transkripčné faktory. Tie v rámci iniciácie transkripcie vykonávajú úlohy, ktoré prokaryotická RNA-polymeráza vykonáva sama: rozoznávanie TATA-boxu, prikladanie RNA-polymerázy II k promótoru, rozpletenie dvojzávitnice DNA a fosforylovanie RNA-polymerázy II, čím je aktivovaná k transkripcii. GC-box a CAAT-box urýchľujú iniciáciu väzbou s ďalšími transkripčnými faktormi. Ďalej už RNA-polymeráza II katalyzuje syntézu RNA bez transkripčných faktorov.
Koniec transkripcie u eukaryot je signalizovaný sekvenciou AATAAA, ktorá sa označuje ako polyadenylačný signál, pretože signalizuje, že po jeho transkripcii bude mRNA v tomto mieste štiepená a post-transkripčne upravená na 3'-konci. Celkovo však eukaryotická mRNA prechádza viacerými post-transkripčnými modifikáciami, a preto sa tento primárny transkript označuje ako pre-mRNA alebo hnRNA (angl. heterogenous nuclear RNA).
Post-transkripčné úpravy link
Hlavný dôvod, prečo musí byť eukaryotická mRNA modifikovaná, je to, že musí byť transportovaná z jadra do cytoplazmy, kde podlieha translácii na ribozómoch. Počas tejto cesty musí byť zabezpečený jej transport cez jadrovú membránu a ochrana pred degradáciou v cytoplazme. Je to teda rozdiel oproti prokaryotickej transkripcii, ktorá je okamžite prepojená s transláciou mRNA.
Eukaryotická pre-mRNA sa upravuje na obidvoch koncoch. Už bolo spomenuté, že 3'-koniec podlieha úpravám, ktoré súvisia s polyadenylačným signálom. Ten spôsobí, že molekula RNA je predĺžená o cca 200-250 nukleotidov adenínu pomocou poly(A)-polymerázy. Táto úprava súvisí predovšetkým s ochranou pred degradáciou z 3'-konca. Úprava 5'-konca spôsobí, že RNA bude správne rozpoznávaná ribozómom a zároveň je to tiež ochrana pred degradáciou cytoplazmatickými enzýmami.

Posledná modifikácia pre-mRNA súvisí so zložením eukaryotických génov. Tie sa totiž skladajú z dvoch typov sekvencií:
- exóny - úseky DNA, ktoré sa prekladajú do sekvencie aminokyselín
- intróny - úseky DNA, ktoré sú z maturovanej mRNA vyštiepené, a teda sa neprekladajú do sekvencie aminokyselín
Táto modifikácia sa nazýva zostrih (angl. splicing - čítaj 'splajsing') intrónov. Znamená to, že tieto sekvencie sa z pre-mRNA enzymaticky vyštiepia a exóny sa spoja dokopy, čím vznikne maturovaná mRNA pripravená na transláciu. V ľudskom genóme má priemerne každý gén až 9 intrónov. Platí, že dlhé gény majú aj dlhé intróny. Extrémnym príkladom je gén pre proteín dystrofín, ktorého pre-mRNA je vyše 2 milióny nukleotidov dlhá, avšak zostrihom 78 intrónov zostane mRNA dĺžky len 14 tisíc nukleotidov!
Intróny sa prakticky nevyskytujú u prokaryot, pretože aj keby tam niekedy boli, akákoľvek náhodná strata úseku obsahujúceho intrón by urýchlila delenie buniek, ktoré by boli v konkurenčnej výhode oproti bunkám s intrónmi. Pre evolučný vznik a vysvetlenie pozoruhodnej hustoty intrónov v dnešnej eukaryotickej DNA existuje viacero hypotéz.