Autor: Peter Pančík
Publikované dňa:
Citácia: PANČÍK, Peter. 2016. Biopedia.sk: Translácia. [cit. 2024-04-23]. Dostupné na internete: <https://biopedia.sk/molekularna-biologia/translacia>.
Translácia predstavuje konečný proces realizácie genetickej informácie štruktúrnych génov. Prebieha na ribozómoch v cytoplazme alebo membránach endoplazmatického retikula za účasti transferových RNA (tRNA), ktoré do ribozómov prinášajú jednotlivé aminokyseliny. Na ribozómoch, ktoré sú akýmisi miniatúrnymi továrňami na výrobu bielkovín, prebieha polymerizácia aminokyselín do súvislého polypeptidového reťazca na základe genetického kódu. Keďže translácia súvisí zo syntézou proteínov, nazýva sa tento proces proteosyntéza.
Ako už bolo povedané, translácie sa zúčastňujú aj transferové a ribozómové RNA, ktorých význam treba pred opisom samotného priebehu translácie vysvetliť.
Štruktúra a význam tRNA link
Transferové RNA sú typom funkčných RNA, ktoré slúžia na prepravu, alebo transfer (odtiaľ ich názov), jednotlivých aminokyselín do ribozómu, kde sa z nich tvorí súvislý polypeptidový reťazec. Každá tRNA predstavuje jednovláknovú RNA dlhú 75-93 báz, ktorá okrem typických RNA nukleotidov obsahuje aj neobvyklé nukleotidy. Tie vznikajú cielenou post-transkripčnou modifikáciou štandardných nukleotidov.
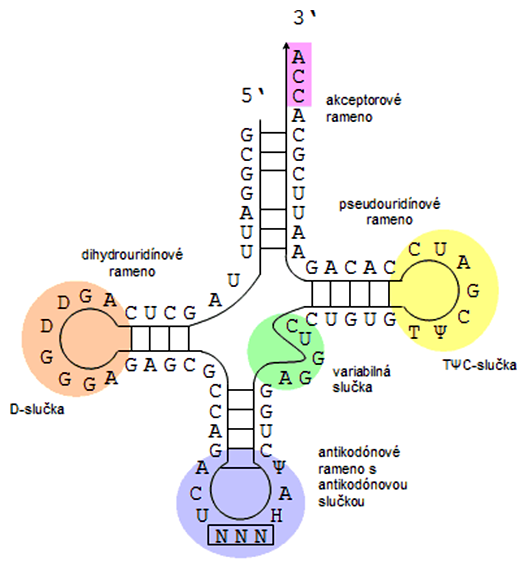
Sekundárna štruktúra tRNA pripomína "ďatelinový štvorlístok". Jednotlivé "lístky" sú zložené z tzv. palindrómov. Palindrómy sú také nukleotidové sekvencie (RNA alebo DNA), ktoré sú symetrické tak, že bázy od osi symetrie smerom doľava sú komplementárne k bázam nachádzajúcim sa vpravo od osi symetrie, napr. 5'-CAGT|ACTG-3'. To spôsobuje na jednovláknovej molekule tvorbu vnútromolekulových spojení, ktoré lokálne tvoria dvojvláknový úsek nazývaný vlásenka (angl. hairpin). Ak sa v strede palindrómu, t.j. v oblasti stredu symetrie nachádzajú nukleotidy, ktoré sa nevedia takýmto spôsobom popárovať, na konci vlásenky sa tvorí jednovláknová slučka (angl. loop). Takýmto prípadom vlásenky so slučkou (angl. hairpin loop) sú aj "lístky" tRNA, ktoré sa označujú ako ramená:
- akceptorové rameno - na jeho koniec sa kovalentne viaže aminokyselina
- antikodónové rameno s antikodónovou slučkou - týmto koncom tRNA rozoznáva genetický kód "zapísaný" v mRNA
- pseudouridínové rameno s TΨC-slučkou - obsahuje modifikovaný uracil - pseudouracil
- dihydrouridínové rameno s D-slučkou- obsahuje modifikovaný uracil - dihydrouracil
- variabilná slučka - môže byť krátka (tRNA typu I) alebo dlhá (tRNA typu II), neovplyvňuje biologickú funkciu tRNA
Obidva konce RNA (3' aj 5') sa nachádzajú v akceptorovom ramene tRNA. Na každú tRNA sa pripája len jedna aminokyselina, a to na posledný adenín 3'-konca tRNA, ktorá končí jednovláknovou prečnievajúcou sekvenciou 5'-CCA-3'. Pre každú z 20 proteínogénnych (tvoriacich proteíny) aminokyselín existuje osobitný enzým - aminoacyl-tRNA-syntetáza, ktorý vytvára kovalentnú väzbu medzi tRNA a jej príslušnou aminokyselinou. Rovnaké aminokyseliny môžu byť spájané s rôznymi tRNA, ale rovnaké tRNA viažu len jednu špecifickú aminokyselinu, čo súvisí s degenerovanosťou genetického kódu.
Všetky tRNA nadobúdajú v živom systéme podobnú terciárnu štruktúru tvaru "Γ", ktorou najlepšie pasujú do vstupných miest na ribozóme. Vnútromolekulové interakcie v tRNA však aj počas takéhoto priestorového usporiadania molekuly zostávajú.
Genetický kód link
Zaradenie aminokyseliny do polypeptidového reťazca je dané pomocou troch za sebou idúcich nukleotidov. Takáto trojica nukleotidov, ktorá kóduje určitú aminokyselinu, sa nazýva kodón. Genetický kód je teda systém pravidiel, podľa ktorých jednotlivé kodóny určujú zaradenie štandardných aminokyselín do polypeptidového reťazca. Za jeho rozlúštenie dostal MARSHALL NIRENBERG (1927-2010) v roku 1968 Nobelovu cenu za fyziológiu alebo medicínu.
Kodónmi sa označujú sekvencie nukleotidov mRNA, ku ktorým sú komplementárne antikodóny na antikodónovom ramene tRNA. Čítanie genetického kódu teda znamená rozpoznávanie kodónov na mRNA pomocou antikodónov na tRNA a prebieha na ribozóme počas proteosyntézy. Prebieha presne od jedného kodónu k druhému, čo znamená, že jednotlivé trojice nukleotidov sa pri čítaní neprekrývajú.
Genetický kód pozostáva zo 64 kodónov (43 = 64). Zmysel kodónu je jeho schopnosť kódovať aminokyselinu. U drvivej väčšiny živých organizmov má genetický kód rovnaký zmysel. Táto vlastnosť genetického kódu sa nazýva univerzálnosť. Od univerzálneho genetického kódu existujú len minimálne odchýlky (u niektorých nižších skupín organizmov a u mitochondrií).
Niektoré kodóny nekódujú žiadnu aminokyselinu, ale majú signalizačnú funkciu. Niekedy sa nazývajú nezmyselné kodóny. Signalizácia je potrebná na vymedzenenie sekvencie mRNA, ktorá bezprostredne súvisí so syntézou proteínov, teda určuje začiatok a koniec syntézy polypeptidového reťazca (pretože nie celá mRNA je prekladaná do sekvencie aminokyselín). Takýto úsek na mRNA, ktorý obsahuje úplnú informáciu pre syntézu polypeptidu, sa nazýva čítací rámec. Začiatok syntézy je určený iniciačným kodónom AUG, ktorý kóduje aminokyselinu metionín (okrem signalizačnej funkcie má teda aj zmysel). Koniec syntézy polypeptidu signalizuje jeden z terminačných kodónov UAA, UAG alebo UGA, ktoré nekódujú žiadnu aminokyselinu.
Čiže kodónov, ktoré kódujú aminokyseliny, je dohromady 61 (64 - 3 terminačné kodóny). Proteinogénnych aminokyselín je však len 20. Z toho vyplýva, že niektoré aminokyseliny (nie všetky ale väčšina) sú kódované rôznymi kodónmi (synonymné kodóny). To je umožnené existenciou rôznych tRNA, ktoré majú odlišné antikodóny ale nesú rovnakú aminokyselinu. Táto vlastnosť genetického kódu sa nazýva degenerovanosť.
V tabuľke genetického kódu sa kodóny, ktoré kódujú tú istú aminokyselinu, odlišujú hlavne v tretej pozícii nukleotidu. Ak sa zmenou 3. nukleotidu v kodóne nezmení jeho zmysel, takáto štvorica kodónov sa nazýva kodónová rodina. Dva synonymné kodóny, ktoré majú na 3. pozícii nukleotidu purín (A,G) alebo pyrimidín (U,C), sa nazývajú dvojkodónová sada.
| 2. báza | ||||
| 1. báza | U | C | A | G |
| U | UUU fenylalanín UUC fenylalanín UUA fenylalanín UUG fenylalanín | UCU serín UCC serín UCA serín UCG serín | UAU tyrozín UAC tyrozín UAA STOP UAG STOP | UGU cysteín UGC cysteín UGA STOP UGG tryptofán |
| C | CUU leucín CUC leucín CUA leucín CUG leucín | CCU prolín CCC prolín CCA prolín CCG prolín | CAU histidín CAC histidín CAA glutamín CAG glutamín | CGU arginín CGC arginín CGA arginín CGG arginín |
| A | AUU izoleucín AUC izoleucín AUA izoleucín AUG START, metionín | ACU treonín ACC treonín ACA treonín ACG treonín | AAU asparagín AAC asparagín AAA lyzín AAG lyzín | AGU serín AGC serín AGA arginín AGG arginín |
| G | GUU valín GUC valín GUA valín GUG valín | GCU alanín GCC alanín GCA alanín GCG alanín | GAU kys. asparágová GAC kys. asparágová GAA kys. glutámová GAG kys. glutámová | GGU glycín GGC glycín GGA glycín GGG glycín |
Genetický kód je teda:
- tripletový
- neprekrývajúci sa
- univerzálny
- degenerovaný
Štruktúra a funkcia ribozómov link
Ribozómy tvoria súčasť každej prokaryotickej a eukaryotickej bunky. Ich úlohou je syntéza proteínov v procese translácie. V priemere majú ~20 nm a skladajú sa zo 65% ribozomálnej RNA (rRNA) a 35% ribozomálnych proteínov. Predstavujú teda komplex ribonukleovej kyseliny a proteínov (ribonukleoproteínový komplex). Ribozómy boli prvýkrát pozorované špeciálnou technikou mikroskopovania v tmavom poli v 30-ich rokoch 20. storočia a neskôr elektrónovým mikroskopom, kedy bola bližšie charakterizovaná ich štruktúra.
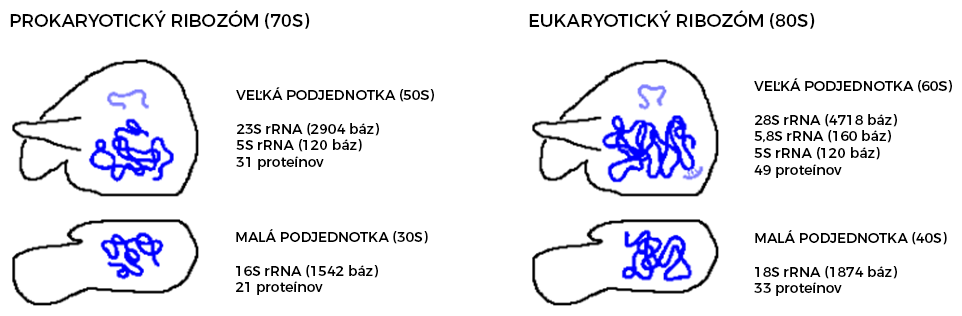
Prokaryotické a eukaryotické ribozómy sa odlišujú vo veľkosti a zložení. Často používaným meradlom hmotnosti ribozómov je sedimentačný koeficient. Udáva sa v jednotkách Svedberg (S), po nositeľovi Nobelovej ceny za chémiu (1926) THEODOROVI SVEDBERGOVI (1884-1971), ktoré určujú rýchlosť sedimentácie častíc pri ultracentrifugácii. Čím väčšie (ťažšie) sú molekuly, tým je väčší sedimentačný koeficient.

Voľné ribozómy sú lokalizované v cytoplazme prokaryotickej a eukaryotickej bunky, kde syntetizujú proteíny. Niektoré proteíny však vyžadujú, aby ich syntéza bola oddelená od cytoplazmy. Patria k nim napr. proteíny bohaté na cysteíny, ktoré tvoria disulfidické väzby, zložené proteíny (napr. glykoproteíny) alebo proteíny slúžiace na sekréciu z bunky. Takéto proteíny veľmi krátko po začiatku svojej syntézy nasmerujú voľné ribozómy na membránu endoplazmatického retikula (ďalej ER), kde už ich syntéza pokračuje do cisterien ER. ER pokryté ribozómami sa nazýva drsné endoplazmatické retikulum.
Ultraštruktúra ribozómov link
Ribozómy pozostávajú z malej a veľkej ribozomálnej podjednotky. U eukaryot dochádza k ich formovaniu už v jadre, odkiaľ sa hotové podjednotky dostávajú cez jadrové póry do cytoplazmy. Za "kľudového" stavu sú oddelené, spájajú sa až pri translácii, po ktorej sa opäť rozchádzajú.
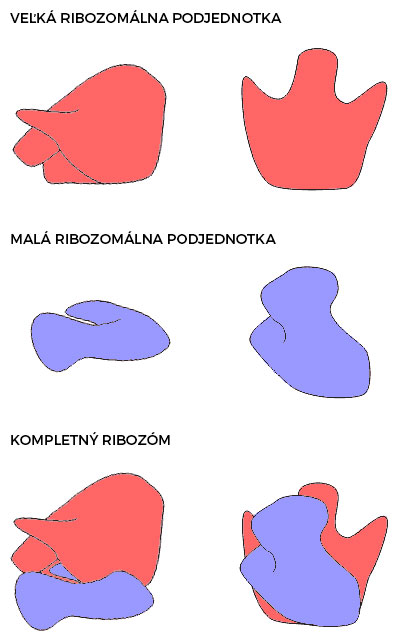
Na ribozóme sú definované viaceré miesta, ktoré interagujú s jednotlivými molekulami (mRNA, tRNA, pomocné proteíny) zúčastňujúcimi sa procesov translácie:
- väzbové miesto pre mRNA - umiestnené na malej podjednotke
- aminoacylové miesto (A miesto) - vstupujú sem tRNA, na ktorých je naviazaná príslušná aminokyselina
- peptidylové miesto (P miesto) - v tomto mieste nastáva predlžovanie polypeptidového reťazca práve o jednu aminokyselinu
- peptidyltransferázové miesto - tvorba peptidovej väzby
- výstupné miesto pre deacylovanú tRNA (E miesto) - tRNA, ktorá odovzdá svoju aminokyselinu, odchádza z ribozómu v tomto mieste
- väzbové miesta pre pomocné translačné proteíny - ďalšie miesta, na ktoré sa viažu pomocné proteíny (tzv. faktory) urýchľujúce a regulujúce proces proteosyntézy
Všeobecný priebeh translácie link
Prokaryotická a eukaryotická translácia prebieha v 3 fázach:
Iniciácia translácie link
Najprv je rozoznávaná tRNA s iniciačnou aminokyselinou, ktorá vytvorí komplex s voľnou malou ribozomálnou podjednotkou a mRNA. Tohto rozpoznávania sa zúčastňujú aj pomocné translačné proteíny, tzv. iniciačné faktory. Tie okrem iného zabezpečia, že iniciačná aminokyselina vstúpi rovno do P miesta, kde sa nachádza iniciačný kodón AUG, tzn. že sa preskočí A miesto. Po vytvorení komplexu dôjde k uvoľneniu iniciačných faktorov a spojeniu malej ribozomálnej podjednotky s veľkou.
Elongácia translácie link
Genetický kód na mRNA sa číta po tripletoch, ktoré sú rozoznávané príslušnými tRNA s naviazanými aminokyselinami. Tieto tRNA vstupujú do A miesta, kde ich nasmeruje jeden z elongačných faktorov. Ak sa antikodón tRNA správne spáruje s kodónom na mRNA, peptidyltransferáza vytvorí peptidovú väzbu medzi novou aminokyselinou a rastúcim polypeptidovým reťazcom. Potom sa ribozóm posunie presne o jeden kodón pozdĺž mRNA (translokácia ribozómu), čo presunie deacylovanú tRNA do E miesta, polypeptid napojený na tRNA do P miesta a A miesto sa uvoľní. Deje sa to za pôsobenia ďalšieho elongačného faktora a spotreby energie (hydrolýza GTP).
Terminácia translácie link
Syntéza proteínu končí, ak ribozóm narazí na jeden z terminačných kodónov (UAA, UAG, UGA). K týmto kodónom nie sú komplementárne antikodóny žiadnych tRNA nesúcich aminokyselinu. Namiesto toho sú rozoznávané terminačnými faktormi, ktoré sa svojou 3D štruktúrou podobajú aminoacyl-tRNA (tzv. molekulárne mimikry). Ich účinkom dôjde k hydrolýze väzby medzi tRNA a polypeptidovým reťazcom a jeho uvoľneniu z ribozómu. Ten sa potom opäť rozpadá na voľné podjednotky, ktoré môžu iniciovať novú transláciu.
Podobne ako v prípade transkripcie, hotový translačný produkt podlieha zmenám, na ktorých konci je proteín schopný vykonávať svoju biologickú funkciu. To zahŕňa odštiepenie alebo modifikáciu iniciačnej aminokyseliny, chemickú modifikáciu ďalších aminokyselín, tvorbu disulfidických mostíkov medzi cysteínovými zvyškami, pripojenie cukorných zvyškov (glykoproteíny) alebo prostetických skupín (napr. hem v hemoglobíne) a tvorbu terciárnej štruktúry.
Prokaryotická translácia link
U Escherichia coli prebieha syntéza polypeptidového reťazca rýchlosťou 10-20 aminokyselín za sekundu. Napodiv je tento proces veľmi presný, pretože frekvencia zaradenia nesprávnej aminokyseliny, často však podobného chemického zloženia, je 1:2000.
U baktérií nastupuje translácia často ešte pred ukončením transkripcie mRNA. Pritom najdôležitejší translačný signál predstavuje tzv. vedúca sekvencia (Shine-Dalgarnova sekvencia) 5'-AGGA-3' umiestnená ešte pred iniciačným kodónom na mRNA. Tá je komplementárna k sekvencii 3'-UCCU-5', ktorá sa nachádza na 3'-konci 16S-rRNA na malej ribozomálnej podjednotke. Prokaryotická mRNA tak už počas svojej syntézy môže byť pokrytá mnohými ribozómami, čo sa označuje pojmom polyzóm. Bezprostredné časové a priestorové prepojenie bakteriálnej transkripcie s transláciou využívajú niektoré regulovateľné transkripčné jednotky.
Typickým znakom bakteriálnej translácie je modifikácia iniciačnej aminokyseliny metionínu na formylmetionín. Pripájanie karbonylovej skupiny na NH2-skupinu metionínu prebieha po naviazaní metionínu na iniciačnú transferovú RNA, s označením tRNAfMet. Takáto tRNAfMet s naviazaným formylmetionínom je rozoznávaná iniciačným faktorom IF2, čo zabezpečuje, že sa formylmetionín viaže na iniciačný kodón AUG a tvorí prvú aminokyselinu v polypeptidovom reťazci. Po zaradení formylmetionínu často nastáva jeho modifikácia (odstránenie karbonylovej skupiny) a až v 50% prípadov dochádza neskôr k odštiepeniu iniciačnej aminokyseliny. Všetky iniciačné faktory (IF1 až IF3) sa viažu len s voľnou malou ribozomálnou podjednotkou a po spojení ribozómu sa z nej uvoľňujú.
Elongácia translácie prebieha podľa všeobecného princípu popísaného vyššie. Čo sa však stane, ak sa vo vnútri čítacieho rámca objaví ďalší iniciačný kodón AUG ? Nič zvláštne. Ten je rozoznávaný transferovou RNA s označením tRNAMet s naviazaným metionínom, ktorý vytvorí komplex s elongačným faktorom EF-Tu, a ten spôsobí, že tRNAMet vstúpi do A miesta podobne ako aj ostatné tRNA pri elongácii. Podobný princíp rozoznávania START kodónu a vnútorných AUG kodónov funguje aj u eukaryot. Translokáciu ribozómu zabezpečuje elongačný faktor EF-G, ktorý hydrolyzuje GTP. Prokaryotickú termináciu translácie riadia terminačné faktory s označením RF1 až RF3.
Prokaryotická translácia je veľmi častým terčom dnešných moderných liečiv, ktoré sa zameriavajú na jej štruktúrne a funkčné špecifiká, pričom nedochádza k ovplyvneniu eukaryotickej translácie ani translácie mitochondriálnej, pretože tá je chránená relatívne bezpečnou bariérou v podobe dvojitej mitochondriálnej membrány.
Eukaryotická translácia link
U eukaryot je translácia oddelená od transkripcie časovo aj priestorovo (transkripcia v jadre, translácia v cytoplazme). Eukaryotickej mRNA chýba Shine-Dalgarnova sekvencia, ktorá je prítomná u prokaryot. Rozlišovacím prvkom je v tomto prípade post-transkripčne modifikovaný 5'-koniec mRNA, ktorý je rozoznávaný eukaryotickými iniciačnými faktormi. Princíp tvorby komplexu mRNA, malej ribozómovej podjednotky a iniciačnej tRNA je rovnaký ako u prokaryot. Po jeho vytvorení dochádza k posúvaniu malej ribozomálnej podjednotky po reťazci mRNA až po iniciačný kodón AUG, kde dôjde k jeho rozoznaniu a spojeniu malej podjednotky ribozómu s veľkou za súčasného uvoľnenia iniciačných faktorov. Tento tzv. scanning model súvisí s tým, že AUG kodón je od 5'-konca mRNA značne vzdialený v porovnaní so vzdialenosťou medzi AUG kodónom a Shine-Dalgarnovou sekvenciou u prokaryot. U eukaryot je iniciačnou aminokyselinou metionín, bez modifikácie.
Eukaryotická elongácia a terminácie translácie je v princípe podobná prokaryotickej.
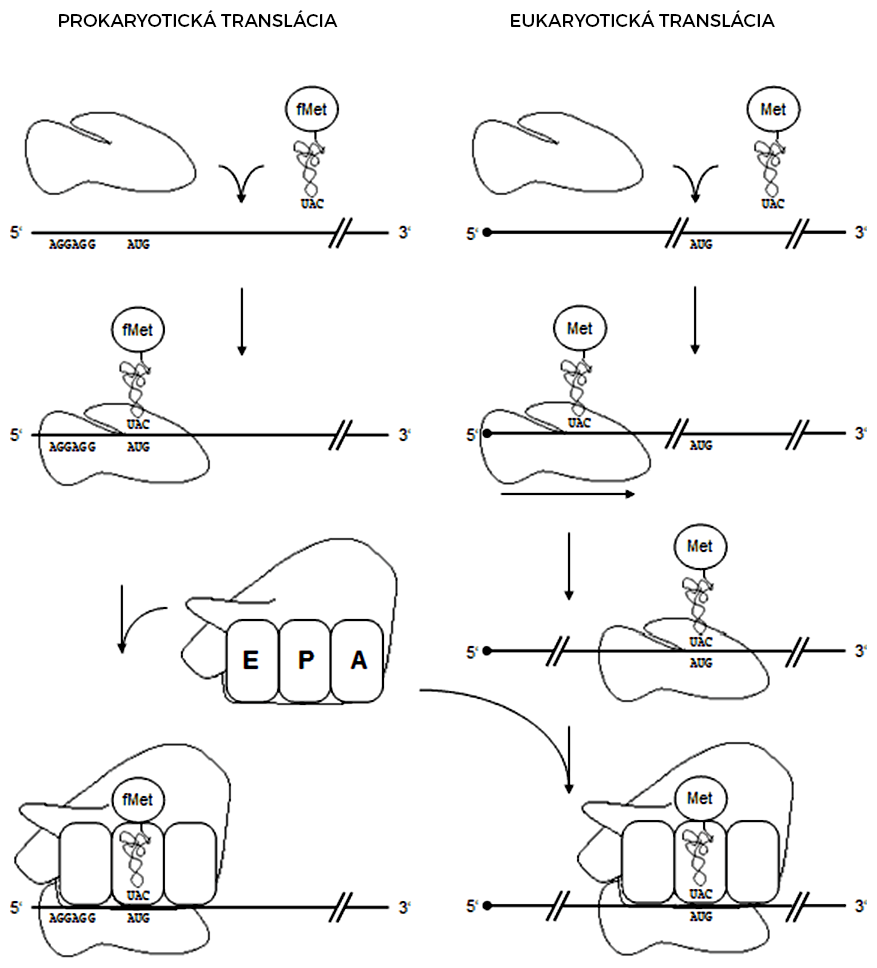
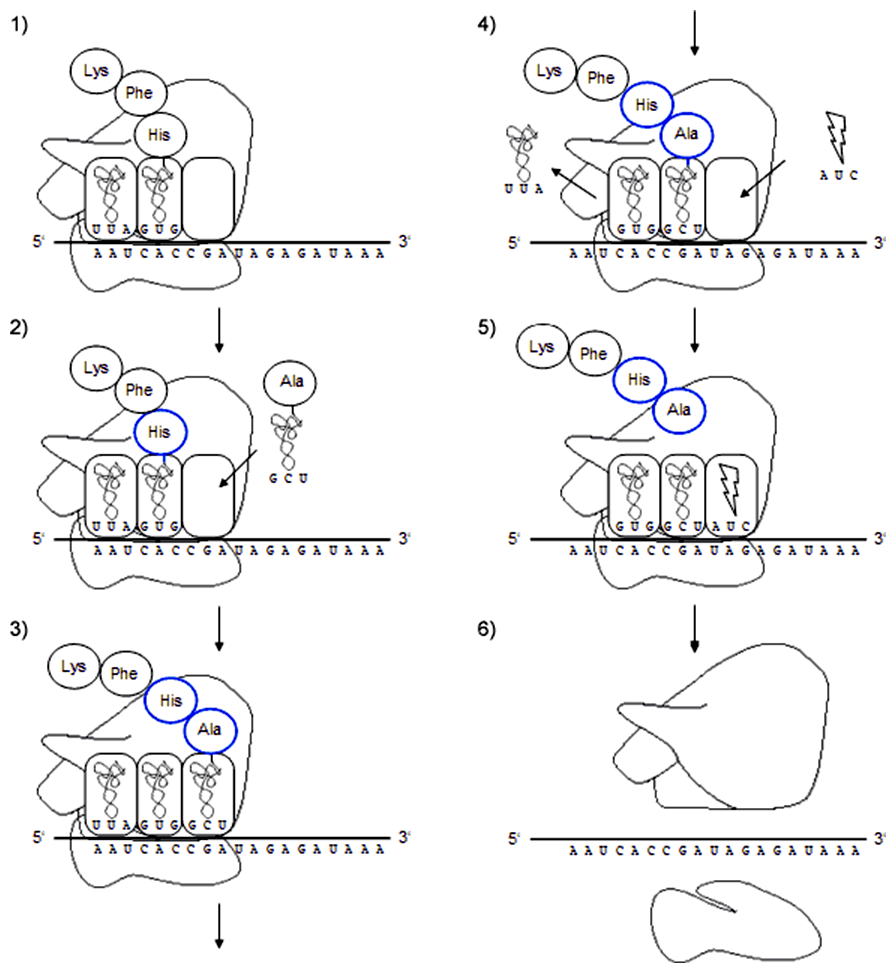
- V E mieste je deacylovaná tRNA (pôvodne s fenylalanínom), v P mieste je peptidyl-tRNA, v A mieste sa čaká na tRNA s príslušným antikodónom.
- Do A miesta prichádza tRNA s alanínom, ktorej antikodón je komplementárny ku kodónu na mRNA.
- Peptidyltransferáza vytvorí peptidovú väzbu medzi peptidovým reťazcom a aminokyselinou v A mieste.
- Translokácia ribozómu. Z E miesta vypadne pôvodná deacylovaná tRNA a jej miesto obsadí nová deacylovaná tRNA, peptidyl-tRNA sa posunie do P miesta a A miesto sa uvoľní. Nasleduje kodón, ku ktorému nie je komplementárny antikodón žiadnej tRNA.
- K STOP kodónu sa naviaže terminačný faktor, ktorý hydrolyzuje väzbu medzi peptidovým reťazcom a tRNA v P mieste.
- Translácia končí vyviazaním všetkých molekúl z mRNA. Ribozóm sa rozpadá na veľkú a malú podjednotku.