
Vzťahy medzi alelami jedného génu link
V bunkách diploidných organizmov je každý gén determinujúci určitý znak zastúpený dvoma alelami, ktoré tvoria alelový pár. Tieto alely sú lokalizované na identických lokusoch párových homologických chromozómov. Ak sú obe alely daného génu zhodné, organizmus sa pre daný znak označuje ako homozygot. Ak je alelový pár tvorený dvoma funkčne alebo štruktúrne odlišnými alelami, ide o heterozygota.
Výsledný fenotypový prejav sledovaného znaku u heterozygota priamo závisí od vnútrogénovej interakcie týchto dvoch odlišných alel. Na základe ich funkčného vzťahu a miery, akou sa spoločne podieľajú na tvorbe fenotypu, rozlišujeme niekoľko základných modifikácií:
- úplná dominancia
- neúplná dominancia
- kodominancia
- superdominancia
Úplná dominancia link
Alelový pár môže byť tvorený dvomi nerovnakými alelami. Takáto konštitúcia alel sa nazýva heterozygotná. Ak je jedna alela svojím účinkom schopná maskovať výsledný prejav druhej alely, hovoríme o vzťahu úplnej dominancie. Dominantná alela sa spravidla označuje veľkým písmenom (A) alebo znakom +. Homozygotná konštitúcia alel AA alebo ++ sa nazýva dominantný homozygot a genotyp aa je tzv. recesívny homozygot.
Príklad: Príkladom úplnej dominancie je tvar semena hrachu: genotypy SS a Ss predstavujú hladký tvar, recesívny homozygot ss má zvráskavené semená.


Vysvetlenie dominantného účinku alely spočíva v tom, že recesívna alela kóduje nefunkčný produkt (najčastejšie enzým), ktorý je však možné úplne nahradiť funkčným proteínom kódovaným druhou (dominantnou) alelou.
Recesívne dedičné ochorenia bývajú často spojené práve s týmto typom dedičnosti. Prejavujú sa len vtedy, ak sa recesívna alela dostane do homozygotného stavu.
Neúplná dominancia link
Ak heterozygot vykazuje fenotyp, ktorý sa nachádza niekde „medzi“, označujeme takýto vzťah medzi alelami za neúplne dominantný a dedičnosť takéhoto znaku sa nazýva neúplná (intermediárna) dedičnosť. Alely takéhoto typu sa spravidla označujú rovnakým písmenom s odlišným indexom, ktorý naznačuje, že sa jedná o menšiu zmenu v porovnaní so vzťahom úplnej dominancie.
Príklad: Príkladom môže byť sfarbenie peria u kura domáceho: homozygot CBCB je sfarbený dočierna, homozygot CWCW je biely a heterozygot CBCW má sivé perie.


Molekulárnym podkladom neúplnej dominancie je tzv. haploinsuficiencia – jav, kedy polovičná génová dávka (jedna alela Cᴮ) nestačí pokryť syntézu dostatočného množstva produktu a výsledkom je len neúplné sfarbenie peria.
Kodominancia link
Špeciálnym typom dominancie je tzv. kodominancia, kedy sa obidve alely správajú ako dominantné, pričom heterozygot vystupuje ako dvojitý homozygot, u ktorého sa fenotypovo prejavujú obidve alely rovnako silným účinkom. Príkladom a zároveň molekulárnym vysvetlením tohto javu je krvný systém MN človeka. Alela Lᴹ syntetizuje antigén M, alela Lᴺ syntetizuje antigén N, pričom heterozygot syntetizuje obidva antigény rovnocenne.
Superdominancia link
Superdominancia je vzťah, kedy má heterozygot (Aa) intenzívnejší (silnejší) fenotypový prejav znaku ako obaja homozygoti, a to vrátane dominantného homozygota (AA).

Génové interakcie link
Predstava individuálnych génov determinujúcich určitý fenotyp je nesprávna a do značnej miery zavádzajúca. Fenotyp je výsledkom komplexnej spoluúčasti mnohých biochemických reakcií a molekulových interakcií, ktoré sú geneticky predurčené viacerými génmi. Postupným odhaľovaním molekulárnej podstaty fenotypu sa odkrýva celá rada ďalších faktorov, medzi ktorými môžeme len ťažko definovať absolútny vzťah dominancie a recesivity.
Génové interakcie sú prejavy dvoch alebo viacerých lokusov, ktoré fenotypovo vystupujú ako jeden znak. Je dôležité zdôrazniť, že génové interakcie sa týkajú kvalitatívnych znakov. Teda aj keď sa na ich prejave podieľa viac génov, stále vytvárajú presne ohraničené fenotypové kategórie (napríklad štyri tvary hrebeňa u kuraťa) a nepodliehajú vplyvu prostredia tak, ako je to pri kvantitatívnych znakoch.
Zistiť sa dajú iba podľa fenotypového prejavu znaku, keď v druhej filiálnej generácii má znak dve, tri alebo štyri fenotypové kategórie s početnosťou, ktorá sa dá odvodiť pôsobením najmenej dvoch génov alebo dvoch alelových párov. Číselné pomery fenotypových kategórií pri interakciách génov veľkého účinku sú rôzne, ale dajú sa odvodiť od štiepneho pomeru dihybridného kríženia \( 9 : 3 : 3 : 1 \).
Pri analýze štiepnych pomerov v nasledujúcich prípadoch génových interakcií neuvažujeme o väzbe génov. Existuje viacero variantov génových interakcií, z ktorých je každý podložený odlišnou molekulárnou podstatou:
Interakcie bez zmeny štiepnych pomerov link
Určitý fenotyp vzniká interakciou dvoch alelových párov, pričom každý alelový pár podmieňuje aj vlastný fenotypový prejav. Spolupôsobením sa vytvorí nový fenotyp, ktorý nie je intermediárneho charakteru. Pri tomto type interakcie nenastávajú zmeny mendelistických štiepnych pomerov.
Príklad: Interakciou dvoch alelových párov R a P je ovládaný tvar hrebeňa kura domáceho. Dominantná vloha R podmieňuje ružicovitý tvar hrebeňa, P hrachovitý. Interakciou oboch dominantných faktorov v genotype sa vytvorí orechovitý hrebeň. Homozygotne recesívny stav oboch génov vedie k vzniku listovitého hrebeňa.


Molekulárny podklad tohto javu nie je presne známy. Môžeme predpokladať, že vznik hrebeňa u recesívneho homozygota (rrpp) je determinovaný ďalšími génmi, pričom v prípade ostatných genotypov nastávajú interakcie medzi produktami týchto „neznámych” génov a alelami R alebo P. V prípade oboch dominantných alel nastávajú interakcie aj medzi týmito produktami navzájom, takže výsledkom je v každom prípade odlišný prejav znaku.
Epistáza link
Epistáza zahŕňa interakciu dvoch génov, z ktorých jeden – epistatický gén – maskuje fenotypový prejav druhého – hypostatického génu. V prípade, že za efekt epistatického génu zodpovedá dominantná alela, jedná sa o dominantnú epistázu. Ak je prejav hypostatického génu obmedzovaný dvoma recesívnymi alelami epistatického génu, jedná sa o recesívnu epistázu. V tomto prípade interakcie génov nevzniká žiadny nový fenotyp.
Dominantná epistáza link
Fenotypový štiepny pomer v F₂ generácii je 12 : 3 : 1.
Príklad: V prípade dominantnej epistázy je dominantná alela jedného génu nadradená nad hypostatický gén. Príkladom dominantnej epistázy je tvorba pigmentov tekvice, ktoré ovplyvňujú gény C a D. Pri recesívnom stave oboch génov vzniká zelené zafarbenie tekvice. Ak je gén D dominantný, zelené farbivo sa rozkladá na žlté. Dominantná alela C však vôbec nedovoľuje vznik zeleného sfarbenia. Gén C je tak epistatickým nad génom D.


Recesívna epistáza link
Fenotypový štiepny pomer v F₂ generácii je 9 : 3 : 4.
Divé myši majú sivú srsť, pretože v ich chlpoch sa striedajú pásiky čiernej a žltej farby. Takýto vzor sa nazýva agouti (čítaj „aguti“) a je typický pre viaceré hlodavce, ktorým slúži ako maskovanie v ich prirodzenom prostredí. Okrem toho existujú domestikované formy, ktoré majú čiernu alebo bielu srsť.
Príklad: Z genetického hľadiska je sfarbenie srsti hlodavcov príkladom recesívnej epistázy, na ktorom sa podieľajú dva gény: dominantná alela C určuje vývoj každej farby srsti (jedince cc sú biele), dominantná alela A určuje aguti vzor v kombinácii so žltou farbou (A-sú agouti, aa sú čierne). Z uvedeného vyplýva, že cc je recesívne epistatický alelový pár nad A-, pretože pri chýbaní akejkoľvek farby nemôže byť alelou A diktovaný farebný vzor. V F2 generácii tak vzniká pomer 9 aguti : 3 čierne : 4 biele.


Komplementarita link
Fenotypový štiepny pomer v F₂ generácii je 9 : 7.
Pri komplementarite fungujú dva alelové páry ako enzýmy, ktoré sú spolu potrebné na syntézu určitého produktu. Ak recesívna alela predstavuje nefunkčný enzým, potrebná je aspoň jedna dominantná alela z každého génu, v opačnom prípade k syntéze enzýmu podmieňujúceho určitý znak nedochádza. Fenotyp je tak redukovaný len na dve triedy.
Príklad: Farba kvetu hrachora (Lathyrus odoratus) je podmienená interakciou dvoch génov. Alela C sa dopĺňa s alelou P a výsledkom je fialová farba kvetu. Recesívny homozygot v niektorom z uvedených génov má biele kvety.


Inhibícia link
Fenotypový štiepny pomer v F₂ generácii je 13 : 3.
Pri tomto type génovej interakcie existujú taktiež len dve fenotypové triedy. V tomto prípade gén inhibítor inhibuje prejav druhého alelového páru. Ten sa prejaví len ak je inhibítor v recesívne homozygotnom stave. Taktiež to možno vysvetliť na molekulárnom základe ako enzymatickú dráhu, kde produkt génu I blokuje syntézu koncového produktu biochemickej dráhy.
Príklad: Inhibícia sa prejavuje napríklad v dedičnosti sfarbenia peria kura domáceho. Dominantná vloha A podmieňuje červené sfarbenie peria. Gén I pôsobí ako inhibítor, ktorý bráni prejavu dominantnej vlohy A a vytvára sa biele sfarbenie peria.


Kompenzácia link
Fenotypový štiepny pomer v F₂ generácii je 10 : 3 : 3.
Zvláštnym a vzácnym prípadom vlohovej interakcie je kompenzácia. Nastáva vtedy, keď sa stretnú dve alely rôznych párov, pôsobiacich na určitý znak protismerne, a to prakticky s rovnako silným účinkom, takže sa ich pôsobenie vzájomne ruší.
Príklad: Zakrivenie struku hrachu podľa švu smerom nadol je podmienené dominantnou alelou V, zakrivenie smerom nahor je podmienené dominantnou alelou D. Prítomnosť aspoň jednej dominantnej alely každého génu alebo homozygotne recesívny stav oboch lokusov zodpovedá za vytvorenie rovných strukov.


Duplicita link
Duplicita zodpovedá za niekoľko od seba odlišných typov interakcií. Spoločným zostáva základný charakter, ktorým sa duplicita odlišuje od komplementarity, že totiž istý efekt je vyvolaný viacerými alelovými pármi, z ktorých každý už sám o sebe je schopný daný efekt spôsobovať. Pre duplicitu sa zvykne používať špecifické označenie alelových párov rovnakými písmenami, ktoré sa od seba odlišujú len číselnými indexmi, napríklad A₁, A₂, A₃... resp. a₁, a₂, a₃... Má to zdôrazniť identitu efektu a rovnocennosť týchto génov.
Rozdiely potom vznikajú v závislosti od toho:
- aký je vzťah medzi alelami:
- vzťah s dominanciou – medzi alelami jedného alelového páru je vzťah dominancie a recesivity
- vzťah bez dominancie – k fenotypovému prejavu prispieva každá dominantná alela, takže je rozdiel medzi Aa (jedna dominantná alela) a AA (dve dominantné alely)
- aký je efekt účinnej alely:
- efekt nekumulatívny – ľubovoľná dominantná alela z každého vlohového páru je schopná podmieniť úplný fenotypový prejav
- efekt kumulatívny – každý alelový pár je typický aditívnym účinkom na celkový prejav znaku
Kombináciou spomenutých účinkov môže vzniknúť niekoľko typov duplicity:
- duplicita nekumulatívna s dominanciou (NsD) – každá dominantná alela je schopná podmieniť úplný prejav znaku (Príklad: sfarbenie pliev ovsa, kde stačí jedna akákoľvek dominantná alela B₁ alebo B₂ na to, aby boli plevy tmavé).
- duplicita kumulatívna s dominanciou (KsD) – účinky génov sa sčítavajú (kumulujú), pričom nezávisí, či je v jednom alelovom páre heterozygot alebo dominantný homozygot (vzťah dominancie).
- duplicita kumulatívna bez dominancie (KbD) – na fenotypovom prejave sa podieľa každá jedna dominantná alela, pričom účinok týchto alel sa sčítava (Príklad: intenzita červeného sfarbenia zrna pšenice, ktorá je priamo úmerná počtu prítomných dominantných alel v genotype).

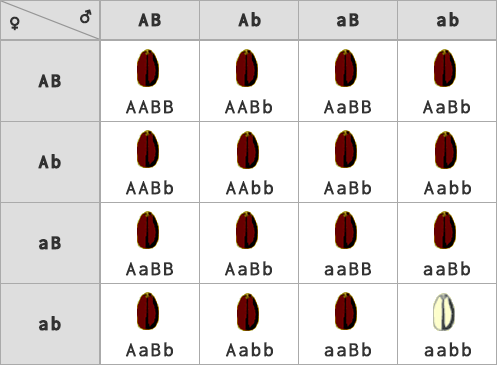




| počet fenotypových tried | typ génovej interakcie | fenotypové štiepne pomery |
| 2 | úplná dominancia | 3 : 1 * |
| 2 | komplementarita | 9 : 7 |
| 2 | inhibícia | 13 : 3 |
| 2 | duplicita nekumulatívna s dominanciou | 15 : 1 |
| 3 | neúplná dominancia | 1 : 2 : 1 * |
| 3 | kodominancia | 1 : 2 : 1 * |
| 3 | dominantná epistáza | 12 : 3 : 1 |
| 3 | recesívna epistáza | 9 : 4 : 3 |
| 3 | kompenzácia | 10 : 3 : 3 |
| 3 | duplicita kumulatívna s dominanciou | 9 : 6 : 1 |
| 4 | interakcie bez zmeny štiepnych pomerov | 9 : 3 : 3 : 1 |
| 5 | duplicita kumulatívna bez dominancie | 1 : 4 : 6 : 4 : 1 |


