
V priebehu evolučnej histórie života na Zemi bol jedným z najzásadnejších zlomov vznik mnohobunkových organizmov. Moderná biológia dnes už takmer s istotou predpokladá, že mnohobunkové živočíchy (Metazoa) vznikli z dávnych bičíkovcov tvoriach kolónie.
Vývoj z jednobunkovca na mnohobunkovca však nebol len jednoduchou otázkou zvyšovania počtu buniek. Zásadným evolučným krokom bola diferenciácia a deľba práce. Bunky v kolónii sa postupne rozdelili na dve funkčné skupiny:
- somatické bunky – telové bunky, ktoré zabezpečovali pohyb či výživu a stratili schopnosť reprodukcie
- generatívne bunky – rozmnožovacie bunky, ktorých jedinou úlohou ostalo rozmnožovanie
Krásnym a dodnes žijúcim modelom takéhoto prechodu je známa zelená riasa váľač gúľavý (Volvox globator), ktorého kolónia už funguje na tomto princípe.
Je dôležité si uvedomiť, že vznik mnohobunkovosti neviedol okamžite k vzniku zložitých tkanív a orgánov. Najprimitívnejšie mnohobunkové živočíchy, ako sú vločkovce (Placozoa) alebo hubky (Porifera), sú len na úrovni zhluku buniek a pravé tkanivá ešte nemajú. Zložité tkanivá a orgánové sústavy sa vyvinuli až oveľa neskôr u evolučne pokročilejších živočíchov, takzvaných epitelovcov (Eumetazoa).
Biogenetický zákon a embryonálny vývin link
ERNST HAECKEL (1866) ako prvý vyslovil hypotézu o vzniku živočíchov z jednobunkovcov a sformuloval biogenetický zákon, podľa ktorého individuálny vývin jedinca (ontogenéza) je skrátené opakovanie historického vývoja jeho druhu (fylogenézy). Ontogenéza tak anatomicky kopíruje fylogenetické štádiá (napríklad zygota zodpovedá jednobunkovcom, gastrula najprimitívnejším mnohobunkovcom).
Skorý embryonálny vývin má tieto etapy:
- oplodnenie – splynutím vajíčka a spermie vzniká diploidná zygota.
- brázdenie – zygota prechádza mitotickými deleniami, výsledkom čoho je zhluk buniek nazývaných blastoméry. Medzi blastomérami sú hlboké ryhy, z čoho pochádza pomenovanie tejto etapy. Celkový objem hmoty sa brázdením zväčša nemení, pretože zygota je chránená od okolitého prostredia pevnou membránou (napríklad u cicavcov je to zona pellucida).
- morula – po niekoľkých deleniach zygoty vzniká plná guľovitá štruktúra, ktorá pripomína plod moruše (z toho názov morula). Fylogeneticky zodpovedá nediferencovaným kolóniám bičíkovcov (napríklad Eudorina). Ryhovanie moruly môže byť nedeterminované (regulačné), kedy bunky ešte nie sú špecializované a teoreticky oddelením každej z nich môže vzniknúť nový jedinec (typické pre druhoústovce). U prvoústovcov sa však stretávame s determinovaným (mozaikovým) ryhovaním, kde je osud každej blastoméry vopred prísne určený a ich flexibilita je značne znížená.
- blastulácia – s ďalším delením sa vo vnútri moruly hromadí tekutina a vytvára sa prvotná telová dutina zvaná blastocél. Plne vyvinutá dutá štruktúra sa nazýva blastula, ktorá je obklopená jednou vrstvou buniek (blastoderm). Fylogeneticky zodpovedá diferencovaným kolóniám (napríklad váľač (Volvox)). U cicavcov blastulu nazývame blastocysta. V nej už dochádza k diferenciácii povrchových buniek zabezpečujúcich uhniezdenie a výživu (trofoblast) a vlastných buniek embrya koncentrovaných na jednom póle (embryoblast).
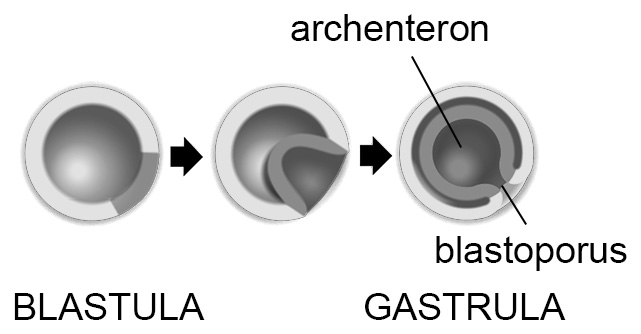
- gastrulácia – kľúčový proces diferenciácie buniek, pri ktorom vznikajú dva zárodočné listy: vonkajší ektoderm a vnútorný endoderm. Výsledným štádiom je gastrula (fylogeneticky zodpovedá stavbe tela primitívnych pŕhlivcov). Dutina, ktorá vo vnútri gastruly vznikne, sa nazýva prvočrevo (archenteron) a do vonkajšieho prostredia ústi otvorom zvaným prvoústa (blastoporus).
Haeckel pri vývine embrya predpokladal, že z dutej blastuly sa jedna stena začala vliačovať dovnútra (invaginácia), čím vznikol hypotetický prapredok všetkých živočíchov, ktorého pomenoval gastrea. I. I. MEČNIKOV (1887) v štúdiu pokračoval a zistil, že tento proces je v skutočnosti z hľadiska fylogenézy vzácny a odvodený jav a popísal iný, pôvodnejší spôsob vzniku vnútornej vrstvy – bunky sa nevliačili ako celok, ale aktívne pohlcovali potravu a vcestovali dovnútra (imigrácia). Tohto predka nazval fagocytela.

Fylogenetické rozdelenie podľa symetrie a zárodočných listov link
Kľúčovým momentom evolúcie bolo vytvorenie zárodočných listov v štádiu gastruly. Predstavujú základné vrstvy, z ktorých sa neskôr vyvinú všetky tkanivá a orgány. Na základe tohto znaku a telesnej symetrie delíme živočíchy do základných vývojových línií.
Živočíchy bez pravých tkanív (Parazoa a Placozoa) link
Z klasického rozdelenia na lúčovité a dvojstranné živočíchy sú vyčlenené najprimitívnejšie skupiny, ktoré netvoria pravé tkanivá a orgány. Patria sem vločkovce (Placozoa), čo sú veľmi jednoduché organizmy bez jasnej symetrie, a hubky (Porifera). Telo hubiek je asymetrické, zriedkavo s náznakmi radiálnej symetrie. Počas ich zárodočného vývinu vznikajú síce dve vrstvy (ektoderm a endoderm), ale dochádza u nich k špecifickej gastrulárnej inverzii, a preto sa u nich nikdy nesformujú skutočné tkanivá.
Lúčovito súmerné živočíchy (Radiata) link
Tieto živočíchy vykazujú radiálnu symetriu, čo znamená, že ich časti tela vybiehajú zo stredovej osi a akýkoľvek rez cez stredovú os vytvorí zrkadlovo súmerné polovice. Sú jednoduchšej stavby a označujeme ich ako dvojlistovce (Diploblastica). Tvoria sa im len dva zárodočné listy: vonkajší ektoderm a vnútorný endoderm. Medzi nimi sa nachádza len nebunková rôsolovitá hmota (mezoglea). Keďže im chýba tretí zárodočný list, nemajú vyvinuté zložité vnútorné orgány ani orgánové sústavy. Patria k nim pŕhlivce a rebrovky.
Dvojstranne súmerné živočíchy (Bilateralia) link
Vykazujú bilaterálnu symetriu. Cez ich telo je možné preložiť len jednu stredovú rovinu, pričom každá strana je zrkadlovým obrazom tej druhej. Tento typ je fylogeneticky spojený s procesom cefalizácie (vznikom hlavy) a vývojom komplexnejších orgánov. Ide o trojlistovce (Triploblastica). Počas embryogenézy u nich totiž vzniká tretí zárodočný list (mezoderm), ktorý sa včleňuje medzi ektoderm a endoderm. Vďaka mezodermu dochádza k vzniku svaloviny, cievnej, vylučovacej a pohlavnej sústavy a vyvíja sa v ňom druhotná telová dutina (coelom). Na rozdiel od skupiny Radiata, dvojstranovce predstavujú platnú a vývojovo opodstatnenú monofyletickú vetvu.
Historický vývoj systému mnohobunkovcov link
Klasifikácia živočíchov prešla viacerými úpravami, ktoré priamo reflektovali nové anatomické a neskôr molekulárne poznatky. Je mimoriadne dôležité tieto systémy historicky oddeliť, aby nedochádzalo k zlučovaniu umelých historických taxónov s modernými fylogenetickými líniami.
1. Tradičný systém (Novák a kol., 1969) link
Tento systém sa opieral o vtedajšie poznatky a v univerzitnej výučbe prežíval pomerne dlho. V tom čase v ňom ešte nebol zaradený kmeň vločkovcov (Placozoa). Ten bol síce objavený už v roku 1883, no ako samostatná skupina živočíchov bol popísaný a uznaný až v 70. rokoch 20. storočia. Živočíchy tento systém delil na štyri oddelenia:
- oddelenie: Morulovce (Mesozoa)
- oddelenie: Hubky (Porifera)
- oddelenie: Mechúrniky (Coelenterata)
- oddelenie: Dvojstranovce (Bilateralia)
2. Prechodný systém (Matis a Vilček 1986, 1987; Majzlan 1998) link
Tento systém reagoval na objav vločkovcov a na fakt, že mechúrniky a dvojstranovce majú k sebe evolučne oveľa bližšie (tvoria pravé tkanivá) než k hubkám či morulovciam. Živočíchy sa stále delili na štyri oddelenia, ale s novou vnútornou štruktúrou:
- oddelenie: Vločkovce (Placozoa)
- oddelenie: Morulovce (Mesozoa)
- oddelenie: Hubkovce (Parazoa, Porifera)
- oddelenie: Epitelovce (Eumetazoa)
- pododdelenie: Mechúrniky (Coelenterata)
- pododdelenie: Dvojstranovce (Bilateralia)
3. Moderný systém (Matis a kol. 2002; Franc 2005) link
Zásadným prelomom bolo zistenie, že primitívne morulovce nie sú na polceste medzi prvokmi a hubkami, ale ide o dvojstranovce (Bilateralia), ktoré sú len extrémne zjednodušené parazitizmom. Stratili preto status samostatného oddelenia a boli presunuté na úplný začiatok bilaterálií. Systém tak získal podobu troch hlavných vývojových stupňov:
- oddelenie: Vločkovce (Placozoa)
- oddelenie: Hubkovce (Parazoa)
- oddelenie: Epitelovce (Eumetazoa)
- pododdelenie: Mechúrniky (Coelenterata)
- pododdelenie: Dvojstranovce (Bilateralia)
- infraoddelenie: Morulovce (Mesozoa)
- infraoddelenie: Acélomáty (Acoelomata) – sem boli odčlenené primitívne bezčrevovce (v minulosti radené medzi ploskulice)
- infraoddelenie: Prvoústovce (Protostomia)
- infraoddelenie: Druhoústovce (Deuterostomia)
4. Súčasný univerzitný didaktický systém (Tirjaková, Vďačný, Kocian 2015) link
Najnovšie univerzitné systémy plne rešpektujú tri vývojové stupne z predchádzajúceho systému, no na základe moderných analýz DNA a prehodnotenia znakov reorganizovali vnútro epitelovcov. Z didaktických dôvodov tento systém zachováva niektoré historické kategórie (ako Radiata), no vo vnútri prvoústovcov prináša obrovskú revolúciu: definitívne sa upustilo od klasického delenia podľa telových dutín (napríklad kategória Pseudocoelomata zanikla) a prvoústovce sa rozdelili na dve hlavné (najväčšie), skutočne príbuzenské fylogenetické línie – špirálovce a zvliekavce (drobnú výnimku tu tvorí len malý prechodný nadkmeň morských štetinoústok).
- vývojový stupeň: Vločkovce (Placozoa)
- kmeň: Plakulovce (Phagocytellozoa)
- vývojový stupeň: Hubkovce (Parazoa)
- kmeň: Hubky (Porifera)
- vývojový stupeň: Epitelovce (Eumetazoa, Epitelozoa)
- 1. skupina: Lúčovite súmerné (Radiata) – nahrádza starý názov Coelenterata
- kmeň: Pŕhlivce (Cnidaria)
- kmeň: Rebrovky (Ctenophora)
- 2. skupina: Dvojstranovce (Bilateralia)
- 1. vývojová vetva: Morulovce (Mesozoa)
- 2. vývojová vetva: Acélomáty (Acoelomata)
- kmeň: Bezčrevovce (Acoelomorpha)
- 3. vývojová vetva: Prvoústovce (Gastroneuralia, Protostomia)
- nadkmeň: Špirálovce (Spiralia) – typické špirálovým brázdením vajíčka a trochofórovou larvou
- kmeň: Ploskavce (Platyhelminthes)
- kmeň: Vírniky (Rotifera)
- kmeň: Mäkkýše (Mollusca)
- kmeň: Obrúčkavce (Annelida)
- nadkmeň: Zvliekavce (Ecdysozoa) – charakteristické zvliekaním zrohovateného povrchu pokožky (kutikuly) pod vplyvom hormónov
- kmeň: Hlístovce (Nematoda)
- kmeň: Pomalky (Tardigrada)
- kmeň: Článkonožce (Arthropoda)
- nadkmeň: Špirálovce (Spiralia) – typické špirálovým brázdením vajíčka a trochofórovou larvou
- 4. vývojová vetva: Druhoústovce (Notoneuralia, Deuterostomia)
- kmeň: Ostnatokožce (Echinodermata)
- kmeň: Polochordáty (Hemichordata)
- kmeň: Chordáty (Chordata)
- 1. skupina: Lúčovite súmerné (Radiata) – nahrádza starý názov Coelenterata


